|
Folia faunistica Slovaca (2009) 14 (8): 47-58 |
|
|
|
|
Ornitocenózy antropicky ovplyvňovaného riečneho ekosystému |
||||
| Bird communities of a river ecosystem influenced by human | ||||
|
Radovan Jambor |
||||
|
Katedra zoológie, Prírodovedecká fakulta, Univerzita Komenského v Bratislave, Mlynská dolina B-1, 842 15 Bratislava 4 [radojambor@gmail.com] |
||||
|
Received: 9.7.2009; Accepted: 28.10.2009; Published: 6.12.2009 |
||||
|
Abstract |
||||
|
Our work engage in research into assessment of factors, which have impact on the river Vah`s ecosystem en rapport with the urban environment. One of the methods, how recognise incidence of factors and biota, is the using of birds as an indicator. Birds are well and easily watching in the terrain and the changes in their behaviour suggest, how much strong is the straight effect of human impact. We can compare results of responsive individuals counting and measurement of the size of recorded factors by correlation analyse. In the period 1997 to september 2005 we recorded 150 species of birds on our model area by the city "Trenčín" (Slovakia). 54 from this count was breeding species. We used the method of belt counting and mapping nest territories. Results, in this event as a species richness, suggest on high ecological quality and they are typical earlier for a spot habitats. During the nest seasons in 2006 and 2007 we recorded altogether 33 breeding species particularly in the riparian willow – poplar wood. Density of the nest territories was 293.2 per 10 ha. Most numerous breeders were Fringilla coelebs (dominance 10.30 %) and Turdus pilaris (9.44 %). Results, in this event as a high density, mean, that the wood is a very important area at the regional scale. They probably suggested also on the "island efect", theory from the island biogeography, when the density of nest territories is higher at the island of fragmented vegetation then the density at the surrounding urban landscape. We tried to assess a "weight" of factors, which have impact on the birds populations on the model area using by the behavioural method. After correlation analyse and the metering a frequency of incidences, we detected that the worst impacts on the birds were (by the characteristics of factors) numbers of fishermen and numbers of speed boats (e.g. canoe). That universal method of behavioural response, which we used first, have some advantages, but also disabilities consequent mainly on limited availability. |
||||
|
Key words |
||||
|
Birds communities, willow – poplar floodplain forest, island effect, human impact, behavioural response |
||||
|
Úvod |
||||
|
Výstavba a sprevádzkovanie sústavy vodných diel
"Vážska kaskáda", vysušovanie a zavážanie mokradí, regulácie vodných tokov, to všetko sa podpísalo pod vážne ekologické zmeny v alúviu rieky Váh v posledných 60-tich rokoch. Ich najviditeľnejším prejavom sa stalo postupné vysychanie inundácie v dôsledku zmien v hĺbke hladiny podzemnej vody. Miznú posledné zvyšky lužných lesov, nastupuje xerotermná a teplomilná vegetácia, šíria sa ruderálne, synantropné a nepôvodné invázne druhy rastlín (Deván 1998,
Jambor & Nuhlíčková 2006).
V alúviu Váhu dnes nájdeme optimálne podmienky pre zachovanie pôvodných biotopov už len na niektorých miestach. Paradoxne sa takéto udržali aj v okolí niektorých vodných diel. Ukážkou toho je napríklad hať Trenčianske Biskupice, ležiaca na strednom toku Váhu, takmer uprostred mesta Trenčín. Bez asanačných zásahov sa tu dodnes zachoval rozsiahlejší porast zaplavovaného mäkkého lužného lesa, lokalita, známa ako Trenčiansky luh. O význame tohto územia ako hniezdiska a úkrytu vodného vtáctva sa vedelo už v 80-tych rokoch (Deván & Májsky 1985) a od 90-tych do súčasnosti bolo o lokalite publikovaných niekoľko ornitologických prác. Vtáčím spoločenstvám Trenčianskeho luhu a širšieho okolia sa venoval Jambor (2003), sumarizoval výsledky monitoringu do roku 2003. Čiastočné spracovanie ornitologických dát priamo zo sledovaného územia nachádzame v práci Jambora (2006). Okrajovo sa dotkli územia aj práce Jambora & Nuhlíčkovej (2006, 2008). Trenčiansky luh a priľahlý úsek rieky Váh sú vzhľadom na svoj blízky kontakt s urbánnym prostredím vhodnou modelovou plochou pre sledovanie prieniku a dôsledkov antropických stresorov v riečnom ekosystéme. Predpokladali sme, že vplyv intenzívnej urbanizácie, rast automobilizmu a nadmerné rekreačné využívanie územia sa prejavia aj na druhovom bohatstve a hniezdnej denzite vtáčích spoločenstiev a behaviorálnych prejavoch jedincov jednotlivých druhov. Vtáky majú veľký bioindikačný potenciál a často sa využívajú pri hodnotení vplyvu rôznych antropických aktivít, ktoré zasahujú do životného prostredia (Riffell et al. 1996, Klein 1993, Lukač & Hršak 2005 atď.). Metódy takéhoto hodnotenia sú však veľmi rôznorodé, líšia sa z hľadiska cieľových druhov, typu ekosystému, krajiny, prístupu, účelu, spôsobu vyhodnotenia atď. Princíp väčšiny z nich je však podobný, založený na porovnaní dvoch území, kde na jednom pôsobia hodnotené antropické vplyvy a na druhom chýbajú, resp. majú odlišnú "veľkosť", frekvenciu výskytu alebo intenzitu. Pre konfrontáciu ornitologických dát z týchto území sa používa široké spektrum rôznych charakteristík, od prezencie a absencie charakteristických vtáčích druhov alebo rôzne veľkého súboru indikátorov, cez druhové bohatstvo, denzitu na jednotku plochy, až po rôzne indexy, v závislosti od typu analýzy. Skupinou veľmi efektívnych, z hľadiska času a interpretácie, ale zároveň menej presných metód, sú behaviorálne, čiže také, ktoré využívajú ako signál negatívneho vplyvu zmeny v etologických prejavoch vtáctva. Väčšina z nich však využíva len čiastkové prejavy, najčastejšie hlasové, prípadne únikovú vzdialenosť. Gutzwiller et al. (1994) sa zaoberali napríklad vplyvom vyrušovania vtáctva na výskyt a konzistenciu tzv. primárneho spevu (t.j. hlasný a ďaleko počuteľný teritoriálny spev) vtákov subalpínskeho pásma. Kontrolné lokality bez vplyvu porovnávali s plochami, na ktorých 1 až 2 hodiny prechádzal 1 dobrovoľník. Zaťaženie mapovaných plôch návštevnosťou preskúmali ešte pred začatím sledovania ornitocenózy. Hodnotili ho 2 stupňami frekvencie vyrušovania (zodpovedajúci jednému alebo dvom vyrušeniam za týždeň) a 2 stupňami váhy vyrušovania (25 % ovplyvnenej plochy a 100 % ovplyvnenej plochy). Každá kruhová plocha mala 1 ha, spolu mal 30 štúdijných plôch. Výsledky autori vyhodnotili analýzou rozptylu. Klein (1993) sledoval dopad rekreácie na vodné vtáky na jednom z tropických ostrovov pri Floride. Počas troch náhodne vybraných dní v časovom intervale 28 dní, 13 x za sebou, registroval počet návštevníkov a behaviorálne prejavy vtákov do vzdialenosti 50 m od hrádze (aby sa reakcie dali rozpoznať). Správanie ľudí zaradil do 5 kategórií, rovnako aj správanie vtákov. Vyrušovanie simuloval experimentálne, napríklad pri kategórii 4 išiel krokom rýchlosťou okolo 0,12 m.s -1, pri kategórii 5 prehrával každú minútu ľudský hlas s tou istou intenzitou. Následne vyhodnotil, aká bola medzi frekvenciou odpovede vtákov a disturbanciami závislosť. Výsledky porovnal už s reálnym hodnotením návštevnosti územia, kedy spolu s 10 dobrovoľníkmi sledoval presný počet ľudí. Juricic et al. (2001) využili pri hodnotení vyrušovania vtáctva "poplachovú" vzdialenosť (resp. vzdialenosť, pri ktorej vtáky vydávajú varovné hlasové signály), čo považuje za o niečo presnejšiu charakteristiku ako únikovú vzdialenosť (resp. vzdialenosť odletu), bežne využívanú pri behaviorálnych metódach hodnotenia faktorov. Tieto vzdialenosti chcel determinovať pre vytýčenie „nárazníkovej“ zóny chráneného územia. Poplachová vzdialenosť bola obvykle väčšia pri veľkých druhoch vtákov, menšia pri drobnejších. Zisťoval tiež, ktoré druhy sú schopné prispôsobiť sa správaniu turistov.Behaviorálna odpoveď vtákov nemusí vždy reflektovať človekom spôsobované disturbancie. Podľa Gill et al. (2001) sa zmeny v správaní vtákov často používajú ako miera vnímavosti voči rušeniu. Podľa nich však neplatí, že by druhy, ktoré sa intenzívnejšie vyhýbajú ľudskej prítomnosti, potrebovali zároveň aj väčšiu ochranu, ako tie, ktoré sú tolerantnejšie voči prítomnosti človeka. Autori odporúčajú, aby boli všetky zmeny v správaní vtákov vystavených antropickým plyvom merané zároveň aj zmenami v kondícii. Z jej práce vyplýva aj isté odporúčanie pre správnu interpretáciu výsledkov. Reakcia vtáctva na disturbantné faktory viac – menej nepoukazuje na potrebu ochrany zasiahnutých druhov, ale na potrebu eliminovať dané faktory aj voči druhom, ktoré sa nejavia ako citlivé voči vplyvu alebo z územia krátkodobo vymizli. Vnímavé druhy môžeme považovať z hľadiska použitia tejto metódy len za indikátory zaťaženia. Burger (1981) porovnaním preferencie rôzne zaťažených lokalít napríklad medzi čajkami, zúbkozobcami a bahniakmi poukázal na to, že prítomnosť človeka a jeho disturbantných aktivít priamo na lokalite nemusí byť kľúčovým faktorom výskytu. Bahniaky sa nevyskytovali napríklad na lokalite, kde nebol žiadny ľudský vplyv a kde ostatné vtáky boli prítomné. Ľudské aktivity boli však zaznamenané v okolí tejto lokality, čo bolo podľa neho príčinou absencie bahniakov. Aj táto práca upozorňuje na možnú chybnú interpretáciu behaviorálnej reakcie vtákov. Použitie týchto metód je väčšinou limitované len na niektoré vplyvy a niektoré taxóny vtákov. Doteraz nebola vytvorená prakticky žiadna univerzálna metóda využívajúca behaviorálnu odpoveď (ako takú, bez ohľadu na typ prejavu), ktorá by bola použiteľná pre všetky druhy a vo všetkých ekosystémoch. Cieľom nášho výskumu bolo sumarizovať výsledky monitoringu ornitocenózy modelového územia v období rokov 1997 až 2007, opísať a porovnať jej kvantitatívne a kvalitatívne charakteristiky v hniezdnej sezóne 2006 a 2007, zistiť aké antropické vplyvy pôsobia na územie a aká bola ich váha, resp. poradie významnosti. Za týmto účelom sme použili a zároveň "otestovali" vlastnú modifikovanú behaviorálnu metódu hodnotenia. |
||||
|
Materál a metódy |
||||
|
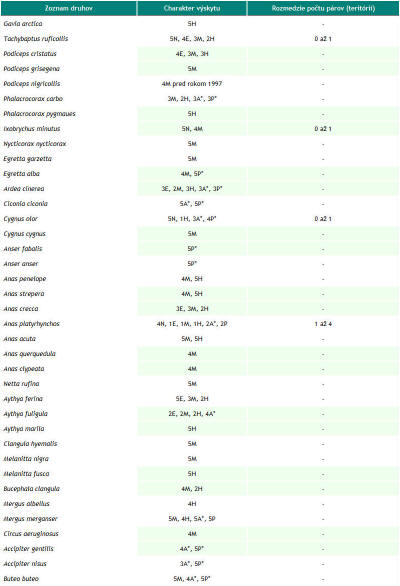
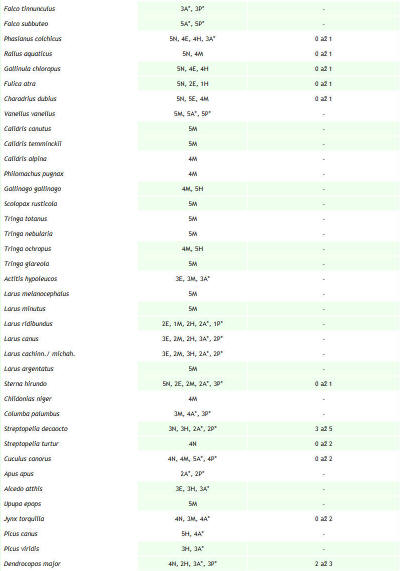
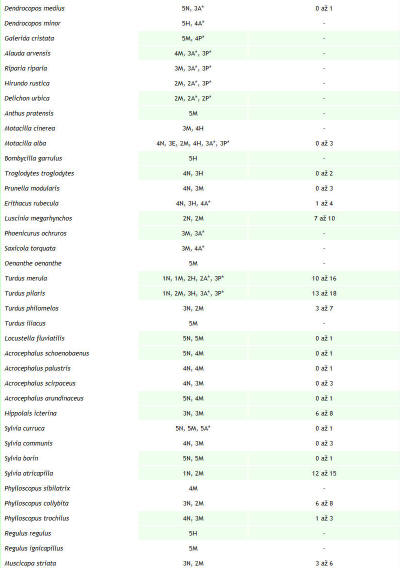
Sledovaná lokalita s rozlohou 32,06 ha leží v Trenčianskej kotline, ktorá je súčasťou orografického celku Považské podolie (490). Prechádza katastrálnym územím obcí Trenčín a na juhu okrajovo Trenčianske Biskupice. Lokalita sa nachádza v štvorci DFS 7174a. Mapovanú plochu tvorí tok rieky Váh, hať Trenčianske Biskupice a Trenčiansky luh. Vytýčená je hrádzou, stavidlami a myslenou čiarou prechádzajúcou korytom niekoľko desiatok metrov južne od cestného mostu. Dĺžka riečneho úseku je 2,5 km, pričom prechádza riečnymi kilometrami medzi 162. až 165. Najväčšie plošné zastúpenie tu majú vodný tok (24,17 ha), zaplavovaný vŕbovo–topoľový lužný les, asi 50–ročný porast asociácie Salici-Populetum (4,40 ha), antropogénne štrkové lavice (1,18 ha) a brehové porasty (0,47 ha). Monitoring vtáčích spoločenstiev Trenčianskeho luhu a priľahlého úseku rieky Váh prebiehal v dvoch fázach. Od roku 1997 do septembra 2005 sme sledovali územie so zameraním najmä na kvalitatívne údaje. Hniezdna početnosť bola iba odhadovaná na základe nepravidelných návštev. Územie bolo sledované po celú dobu zároveň v rámci programu zimného sčítania vodného vtáctva. S presnou kvantifikáciou hniezdnej (počas sezóny v roku 2006) a mimohniezdnej početnosti pásovou metódou (Janda & Řepa 1986) sme začali od septembra 2005. Hniezdnu zložku ornitocenózy v roku 2007 sme zisťovali už metódou mapovania hniezdnych teritórií doplnenou priamym vyhľadávaním hniezd (Janda & Řepa, l.c.). Do schématických máp sme pritom zaznamenávali pozície vtákov pomocou systému štandardných symbolov (Kropil 1992). Vtáky sme registrovali v teréne opticky aj akusticky. Spolu sme od septembra 2005 do októbra 2007 vykonali 110 návštev, a to prevažne v skorých ranných hodinách a za priaznivého počasia. Zistenú abundanciu hniezdnych párov sme v prípade porastu lužného lesa prepočítali na jednotku plochy 10 ha. Výsledky z hniezdnej sezóny v roku 2006 a 2007 (pozri obr. 1) sme porovnali s cieľom zistiť, aký je trend vývoja populácie nidifikujúcich druhov, prípadne aký je rozdiel v presnosti použitých mapovacích metód. Použili sme pritom hodnoty abundancie a denzity hniezdičov, dominancie, druhového bohatstva ornitocenózy (ako počet druhov), Sörensenovho indexu podobnosti, Shannon – Wienerovho indexu diverzity (s prirodzeným logaritmom) a Engenovho indexu equitability (Janda & Řepa 1986). Druhy hniezdiace v poraste lužného lesa sme zaradili do tried dominancie podľa Pelikána (1993). Na doplnenie charakteristiky ornitocenózy územia sme použili rozdelenie vtákov podľa ich hniezdnych gíld (Hudec & Černý 1977, Hudec 1983, 1994). Na základe výsledkov monitoringu ornitocenózy územia sme vytvorili zoznam druhov zaznamenaných v rokoch 1997 až 2007 (pozri Tab. 1). Charakter a dobu výskytu vtáctva uvádzame podľa nasledovných kategórií upravených a doplnených podľa práce Danka et al. (2002): nidifikácia a estivácia (obdobie od apríla do júla), migrácia (marec a august až november), hibernácia (december až február), zálety z blízkeho okolia a preletovanie bez topickej alebo trofickej väzby k územiu. Systém vtákov uvádzame podľa Voousa (1973, 1977). K jednotlivým typom nehniezdneho charakteru výskytu sme priradili stupeň frekvencie výskytu počas celého obdobia sledovania územia. Pre nehniezdiace druhy sú to stupne: 1 – veľmi častý (záznam počas 41 a viac návštev), 2 – častý (počas 40 až 21 návštev), 3 – menej častý (20 až 6), 4 – vzácny (5 až 3) a 5 – raritný (2 až 1). Ku hniezdiacim druhom sme priradili stupeň hojnosti založený na maximálnej zistenej abundancii hniezdnych párov (resp. teritórií) od roku 1997 do roku 2007: 1 – veľmi početný (15 a viac párov), 2 – početný (14 až 9 párov), 3 – menej početný (8 až 4 páry), 4 – málo početný (3 až 2 páry), 5 – nepočetný (1 pár). Odhad hniezdnej početnosti uvádzame v zozname ako rozmedzie zaznamenaného počtu párov alebo teritórií za celé obdobie (1997–2007) a to na základe odhadov, ako aj výsledkov mapovania kvantitatívnymi metódami v rokoch 2006 a 2007. Na území sme registrovali prítomnosť pôsobiacich antropických faktorov a ich veľkosť sme „merali“ prostredníctvom rôznych charakteristík (napr. počet rybárov, počet preletov lietadiel, počet motorových člnov atď.). Mieru reakcie vtáctva sme určili ako pomer reagujúcich jedincov (bez ohľadu na spôsob reakcie, resp. konkrétny etologický prejav) ku všetkým zaznamenaným pre jednotlivé druhy počas návštevy. Intenzitu prejavu vyjadroval samotný počet reagujúcich jedincov. Pre každú návštevu sme vypočítali priemer miery reakcie pre jeden druh. Medzi touto hodnotou a veľkosťou zaznamenného faktora, z ktorých väčšina bola disturbantného charakteru s priamym krátkodobým optickým alebo akustickým prejavom, sme hladali závislosť prostredníctvom korelačnej analýzy. Použili sme pritom Pearsonov korelačný koeficient a jeho štatistickú významnosť sme testovali dvojstranným t-testom (Šmelko 1991). Hodnotu t, porovnávanú s kritickými hodnotami rozdelenia t-Studenta na hladine α = 0,05, sme vypočítali podľa upraveného vzťahu t = |rxy|/ √((n-2)/ (1-rxy2)) (Degma in verb.) pri stupňoch voľnosti 2. Výslednú lineárnu závislosť sme charakterizovali ako silnú priamu (ak r je z inetervalu <0,7; 1>), silnú nepriamu, <-1; -0,7>, slabú priamu alebo nepriamu <-0,5; 0,5> a ak sa r blížilo 0, sledované znaky boli nezávislé (Gregorová & Filová 2004). Výslednú váhu antropických faktorov, ako poradie ich významnosti vzhľadom k ich vplyvu na vtáky, sme určili na základe súčinu štatisticky významného korelačného koeficientu a frekvencie výskytu daného faktora. Zahrnutím frekvencie sme chceli umelo zvýšiť váhu tých faktorov, ktoré sa vyskytujú častejšie, pretože predpokladáme, že opakované pôsobenie bude mať na miestnu ornitocenózu negatívnejší dopad. Synergické a kumulatívne účinky sme nehodnotili, čo mohlo v prípade niektorých faktorov spôsobiť skreslenie. |
||||
|
Výsledky |
||||
|
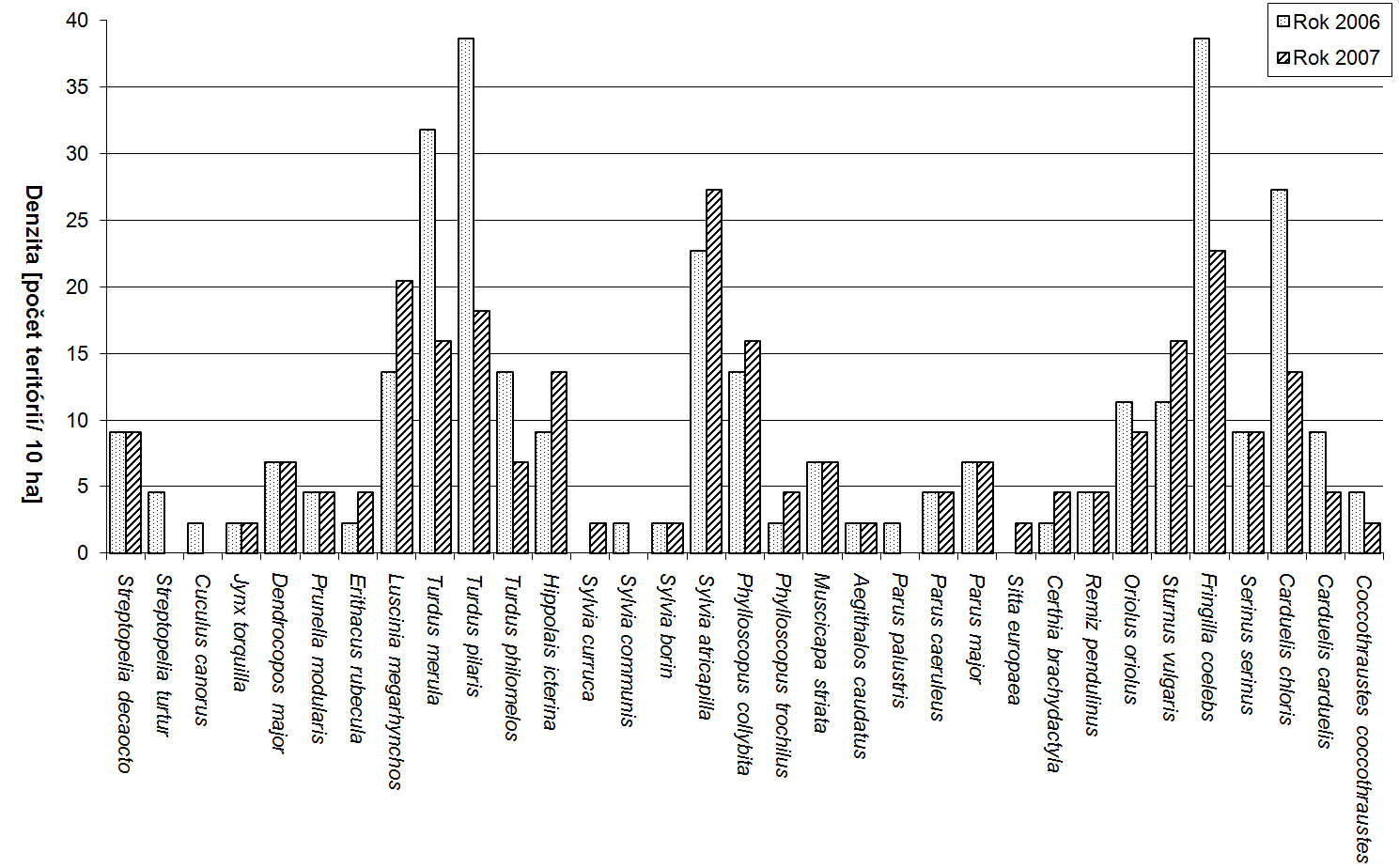
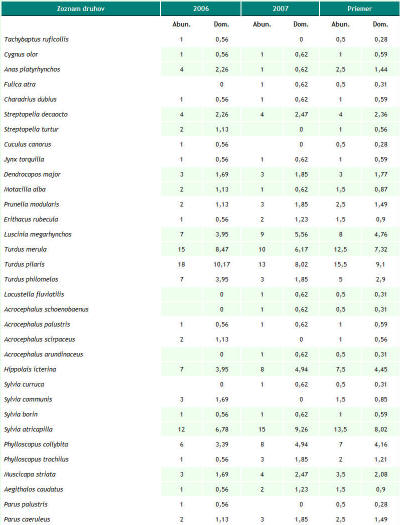
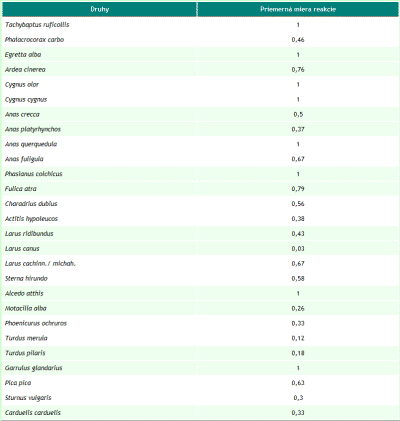
Za obdobie rokov 1997 až 2007 sme na území Trenčianskeho luhu a priľahlého úseku rieky Váh zaznamenali spolu 149 druhov vtákov, z toho 54 hniezdičov. V období od septembra 2005 do októbra 2007, kedy bola venovaná územiu väčšia pozornosť, to bolo 134 druhov a 46 hniezdičov. V priebehu jednotlivých návštev sme pri dodržaní zvolených mapovacích metód registrovali od 10 do 44 druhov (priemerne 28,71). Minimum bolo 22.1.2006 a maximum sme zaznamenali dňa 20.5.2006. Na 1 návštevu územia pripadalo priemerne 316,22 zaznamenaných jedincov vtákov. Najviac, 1137, sme zaznamenali 2.12.2005, z tohto počtu pripadalo až 1018 jedidncov druhu Anas platyrhynchos. Obidva roky (2006 a 2007) majú 98 druhov spoločných, index podobnosti je 94,23 %. Podľa charakteru výskytu sme zaznamenali v období od roku 1997 do roku 2007 na území aj estivujúce (19), migrujúce (108) a zimujúce druhy (59) a druhy zaletujúce z blízkeho okolia (62) a preletujúce bez trofickej alebo topickej väzby k územiu (48) (pozri tab. 1). Jednotlivým druhom pripadá vo väčšine prípadov viacero z týchto kategórií. K hniezdičom (54) sme neprirátali 9 druhov, u ktorých nidifikácia bola len pravdepodobná. Podľa rozdelenia vtákov do hniezdnych gíld sme zaznamenali na území 6 hniezdičov na zemi (Z), 3 hniezdiče v bylinnej etáži (Bz), 3 v bylinnej etáži nad povrchom vodnej hladiny (Bv), 1 na plávajúcich formáciách (P), 9 v krovinách (Kr), 13 v korunách (Ko) a 11 v stromových dutinách (D). Hniezdnu ornitocenózu záujmového územia tvorilo v roku 2006 41 druhov (tvoriacich spolu 177 hniezdnych teritórií, resp. párov), v roku 2007 39 (162 teritórií). Spoločných druhov bolo 34, index podobnosti medzi oboma sezónami je 85%. Prehľad abundancie a dominancie hniezdičov na celom území v rokoch 2006 a 2007 podáva tab. 2. V samotnom poraste lužného lesa (4,4 ha) sme zaznamenali 33 hniezdičov, 31 v roku 2006 (142 hn. teritórií) a 29 v roku 2007 (116 hn. teritórií). Priemerná denzita hniezdnej ornitocenózy bola 293,20 teritórií na 10 ha. Zastúpenie druhov v triedach dominancie bolo nasledovné: 1 druh bol eudominantný, Fringilla coelebs (dominancia 10,30 %; denzita 30,69 ter. na 10 ha), 6 druhov bolo dominantných, Luscinia megarhynchos (6 %; 17,05 na 10 ha), Turdus merula (7,95 %; 23,87 na 10 ha), Turdus pilaris (9,44 %, 28,41 na 10 ha), Sylvia atricapilla (8,69 %; 25,00 na 10 ha), Phylloscopus collybita (5,13 %, 14,78 na 10 ha) a Carduelis chloris (6,81 %, 20,46 na 10 ha), 10 druhov bolo subdominantných, 7 recedentných a 9 subrecedentných. Výsledky z oboch hniezdnych sezón (2006 a 2007) z porastu lužného lesa sme vzájomne porovnali. Spoločných hniezdičov bolo 27, index podobnosti bol 90 %. V roku 2006 boli 2 druhy eudominantné, Fringilla coelebs (11,97 %) a Turdus pilaris (11,97 %), 3 dominantné, 12 subdominantných, 5 recedentných a 9 subrecedentných. V roku 2007 bol 1 druh eudominantný, Sylvia atricapilla (10,34 %), 8 druhov bolo dominantných, 7 subdominantných, 7 recedentných a 6 subrecedentných. V roku 2006 bol index diverzity 3,03 a index equitability mal hodnotu 0,89. V roku 2007 bol index diverzity 3,11 a index equitability 0,92. Prehľad zistených denzít hniezdičov v jednotlivých rokoch podáva obr. 1. Na záujmovom území sme od septembra 2005 do júna 2007 zaznamenali pôsobenie 32 antropických vplyvov, ktoré sme opísali prostredníctvom 69 charakteristík. Najväčšiu frekvenciu výskytu mali charakteristiky: počet návštevníkov na území (0,78 krát na 1 návštevu), počet cyklistov, korčuliarov a kočíkov (0,42 na 1 návštevu), počet rybárov (0,42 na 1 návštevu), počet na území odstavených dvojstopových motorových vozidiel (0,39 na 1 návštevu) a počet pohybujúcich sa nákladných dvojstopových motorových vozidiel do 50 m od územia (0,32 na 1 návštevu). Spolu sme zaznamenali 27 druhov, ktorých jedince reagovali na pôsobiace antropické faktory. Najväčší počet reagujúcich jedincov, 124, bol dňa 20.7.2006, z tohto najviac patrilo druhom Sturnus vulgaris (81) a Anas platyrhynchos (40). Za hodnotené obdobie bola najväčšia priemerná miera rekcie (1, čiže 100% reagujúcich jedincov) zaznamenaná pri 8 druhoch: Tachybaptus ruficollis, Egretta alba, Cygnus olor, Cygnus cygnus, Anas querquedula, Phasianus colchicus, Alcedo atthis a Garrulus glandarius. Naopak najmenšia priemerná miera reakcie bola u Larus canus (0,03) a Turdus merula (0,12). Prehľad zaznamenaných reagujúcich druhov prináša tab. 3. Spôsob reakcie jedincov nebol hodnotený, ale prevažná väčšina reagovala odletom z územia (napr. Anas platyrhynchos, Ardea cinerea), preletom na menšiu alebo väčšiu vzdialenosť (napr. Anas crecca, Phalacrocorax carbo), únikom iným spôsobom (napr. Fulica atra, Tachybaptus ruficollis), varovnými hlasovými signálmi (napr. Anas platyrhynchos, Garrulus glandarius) a rozrušeným správaním (napr. Anas platyrhynchos, Cygnus olor). Korelačná analýza ukázala, že slabá priama závislosť (so štatisticky významným korelačným koeficientom) existuje medzi mierou reakcie a týmito faktormi, resp. ich charakteristikami: počet rybárov (korelačný koeficient 0,21; frekvencia výskytu vplyvu 0,42), počet nízkych preletov (korel. koef. 0,26; frekvencia 0,18), počet rýchlych člnov, ako sú napr. kanoe, (0,41; frekvencia 0,14), počet motorových člnov (0,22; frekvencia 0,03), počte striel mimo územia (0,21; frekvencia 0,09) a počet hlučných (používajúcich zvukové výstražné zariadenia) osobných motorových dvojstopových vozidiel mimo územia (0,21; frekvencia 0,1).
|
||||
|
Diskusia |
||||
|
Záujmové územie sme sledovali takmer 10 rokov, ale až posledné dva sme venovali väčšiu pozornosť aj denzite hniezdnej ornitocenózy a pôsobeniu antropických faktorov. Za toto kratšie obdobie sa nám nepodarilo zachytiť dlhodobo pôsobiace a nepriamo sa prejavujúce vplyvy, ako sú napríklad zmeny v zložení a v štruktúre vegetácie a zmeny v potravnej ponuke. Navzdory tomu však výsledky priniesli aspoň „hrubú“ predstavu o význame územia a jeho zaťažení ľudskými aktivitami. Vysoký počet zaznamenaných druhov je typický skôr pre "bodové" lokality, tvorené menšími akvatickými krajinnými prvkami (napr. štrkoviská, rybníky, vodné nádrže) alebo „ostrovy“ kvalitných hniezdnych a potravných podmienok (napr. fragmenty lesných porastov). Na území sme zaznamenali tiež pomerne vysokú hniezdnu početnosť. Denzita viacerých hniezdičov lužného lesa je nadpriemerná oproti iným lesným ekosystémom aj oproti iným územiam s lužným lesom (pozri tab. 4). Na záujmovom území sme v porovnaní s inými úsekmi rieky Váh zaznamenali väčšiu koncentráciu zimujúceho vodného vtáctva, zapríčinenú ale hlavne vysokým podielom Anas platyrhynchos. Druh sa vyznačuje vyššou adaptibilitou na rušivé vplyvy, v zime tu dokonca prítomnosť človeka využíva (Jambor & Nuhlíčková 2008). Veľký počet zaznamenaných migrujúcich druhov vtáctva na území vyplýva z geomorfologických podmienok Považského podolia. Územie sa nachádza v nadregionálnom migračnom biokoridore (Krumpolec 2004) a poskytuje, okrem pokojnejšej rozšírenej a hlbšej vodnej hladiny, aj plochy s vhodným vegetačným krytom a s odpočinkovými možnosťami (brehové porasty, zárasty pálky a tŕstia, debris). Vysoký počet zaznamenaných druhov, ale aj veľký počet hniezdičov a hniezdnych párov na území dokazuje jeho mimoriadnu ornitologickú a ekosozologickú hodnotu minimálne na regionálnej úrovni. Územie je so svojim poloprírodným lužným lesom v kombinácii so špecifickými hydrologickými pomienkami vodného toku (hať, pravidelné záplavy) ojedinelé. Zároveň však táto zvýšená koncentrácia vtáctva vypovedá aj o tom, že okolitá krajinná matrica je narušená ľudskými aktivitami a má, keďže ide o urbánne prostredie, nižšií stupeň ekologickej stability. Vzniká tu akýsi "ostrovný efekt", kde naše územie predstavuje, bez ohľadu na antropickú záťaž, útočisko pre biotu pôvodného riečneho ekosystému. Väčšinou k takémuto javu dochádza vplyvom fragmentácie pôvodného lesného spoločenstva (Begon et al. 1996). Adaptáciou hniezdiaceho vtáctva na takéto podmienky je potom napríklad zmenšovanie veľkosti, respektíve prekrývanie, hniezdnych teritórií (napr. Mazerolle & Hobson 2004). Dôsledkom je zvýšenie denzity hniezdičov. V našom prípade tento „ostrov“ nevznikol vplyvom fragmentácie väčšieho lesného celku, ale vplyvom samovoľného uchytenia a izolácie porastu už v narušenej a človekom pozmenej krajine. Veľmi vysoké hodnoty denzít hniezdiacich druhov vtáctva boli zaznamenané, pre porovnanie, často práve v prostredí izolovanej mestskej zelene a parkoch v mestách (Danko et al. 2002). Významný vplyv na výsledky výskumu môže mať aj okrajový efekt (Odum 1977, Janda & Řepa 1986). Keďže rozloha nášho porastu lužného lesa je malá (4,4 ha), podiel okraja a jeho účinkov na ploche bude značný, a to aj cez to, že sme sa snažili túto časť porastu z mapovania vylúčiť. Hniezdnu ornitocenózu lužného lesa v roku 2006 a 2007 sme navzájom porovnali. Hodnoty sa v prípade počtu druhov výraznejšie nelíšia, väčší rozdiel je ale v celkovom počte zaznamenaných hniezdnych teritórií (142 a 116). Z hľadiska dominancie bol v oboch prípadoch prevažujúci počet subdominantných druhov. V roku 2006 bol len 1 druh eudominantný, v roku 2007 2 druhy. Indexy diverzity a equitability sa výraznejšie nelíšia, čiže spoločenstvo bolo relatívne vyrovnané. Významný rozdiel je však v hodnotách zaznamenaných denzít hniezdnych teritórií (pozri obr. 1). Rozdiel ale nevypovedá o klesajúcom trende populácií druhov, ale je skôr dôsledkom použitia odlišných kvantitatívnych metód, pásovej v roku 2006 a metódy mapovania hniezdnych teritórií v roku 2007, ktorá je považovaná za presnejšiu. Mestské prostredie zaťažené antropickými stresovými javmi spôsobuje vplyvom postupujúcej urbanizácie ich čoraz častejší prienik na záujmové územie. Ich váhu pôsobenia sme na ploche stanovili zaznamenávaním miery reakcie vtáctva a následnou korelačnou analýzou. Najväčšiu váhu mal zo zaznamenaných antropických faktorov športový rybolov (hodnotená charakteristika "počet rybárov"; váha 0,09), nasledovali vodné športy (charakteristika "počet rýchlych člnov (kanoe a pod.)"; váha 0,06), prelety ľahkých lietadiel (charakteristika "počet nízkych preletov (do 100 m) ľahkých motorových lietadiel"; váha 0,05), prítomnosť a pohyb motorových dvojstopových vozidiel (charakteristika "počet hlučných dvojstopových vozidiel mimo územia (do 1 km)"; váha 0,02) a strelba (charakteristika "počet striel alebo iných hlučných rán mimo územia"; váha 0,02) a opäť vodné športy, zastúpené charakteristikou "počet motorových člnov" (váha 0,01). Keďže nami upravená metóda behaviorálnej odpovede vtáctva nebola ešte mimo našej plochy nikde použitá, nie je možné tieto výsledky porovnávať s literatúrou. |
||||
|
Závery |
||||
|
Od roku 1997 venujeme pozornosť územiu, ktoré svojou ornitocenózou dosahuje kvalitu prirodzených riečnych ekosystémov. Svojim spôsobom (štruktúra, diverzita a kompozícia biotopov) reprezentuje stredný tok Váhu, aký tu bol v období pred osídlením a intenzívnym využívaním človekom. Zároveň nám jeho umiestnenie v úzkom kontakte s mestským prostredím poskytuje príležitosť sledovať, ako postupujúca urbanizácia a čoraz intenzívnejšie využívanie krajiny ovplyvňujú riečny ekosystém. Na území Trenčianskeho luhu a priľahlého úseku rieky Váh sme zhruba za desaťročné obdobie zaznamenali 149 druhov vtáctva (150 po zarátaní starších dát), z toho 54 hniezdičov. Tento počet je pomerne vysoký, typický skôr pre akvatické ekosystému „bodového“ charakteru. Denzity hniezdnych teritórií zaznamenané v poraste lužného lesa sú nadpriemerné. Spolu s počtom druhov poukazujú na mimoriadnu hodnotu územia minimálne na regionálnej úrovni. Výsledky môžu byť čiastočne skreslené malou rozlohou štúdijnej plochy a veľkým zastúpením okraja. Vysoká hustota zaznamenaných teritórií vypovedá pravdepodobne aj o určitom „ostrovnom efekte“, keďže okolitú krajinnú matricu tvorí urbánne prostredie s nižšou ekologickou stabilitou. Vybrané charakteristiky ornitocenózy lužného lesa zistené počas hniezdnych sezón v rokoch 2006 a 2007 sme navzájom porovnali. Rozdiely neboli veľké, týkali sa skôr počtu hniezdnych teritórií a sú dôsledkom použitia rozdielnych mapovacích metód, resp. doplnením pásovej metódy v roku 2007 metódou mapovania hniezdnych teritórií. Na území sme sledovali frekvenciu výskytu a váhu pôsobenia antropických faktorov na vtáky. Pre krátkosť času sme však nevenovali pozornosť dlhodobo sa prejavujúcim a nepriamym faktorom, ako sú napríklad zmeny v štruktúre vegetácie. Veľkosť vplyvov sme merali prostredníctvom charakteristík zodpovedajúcich prevažne počtu alebo pohybu sledovaných objektov a miera reakcie vtáctva na ne zodpovedala pomeru reagujúcich jedincov ku všetkým zaznamenaným počas návštevy. Najčastejšie zistené charakteristiky boli počet návštevníkov na území a počet cyklistov, korčuliarov a kočíkov, ktoré zodpovedajú hodnoteným faktorom „prítomnosť a pohyb ľudí“ a „cyklistika, korčuľovanie a návštevnosť s kočíkmi“. Najväčšiu váhu vplyvu na vtáky mali športový rybolov, vodné športy, prelety ľahkých lietadiel nad územím, prítomnosť a pohyb motorových vozidiel a streľba. "Otestovaním" tejto relatívne univerzálnej metódy behaviorálnej odpovede vtáctva sme dospeli k niekoľkým záverom a odporúčaniam. Jej použitie je vhodné najmä v prípadoch, kedy nemáme dostatok času pre stanovenie antropického zaťaženia určitého územia. Stačí na to prakticky jeden celý rok, prípadne len jedna hniezdna sezóna. Zároveň nie je nevyhnutne nutné mať referenčnú lokalitu bez pôsobenia antropických stresorov. Podstatnou nevýhodou metódy je však jej obmedzená použiteľnosť, nemožno ňou hodnotiť všetky typy antropických stresorov, len také, ktoré je možné zachytiť opticky alebo akusticky v priebehu kratšieho času, také ktoré sa prejavujú priamo a majú disruptívny a disturbantný charakter (zaťaženie rekreačnými aktivitami, rybolovom, leteckou a automobilovou dopravou a pod). Hodnotenie niektorých stresorov je nutné doplniť tiež použitím ďalších metód, obzvlášť v prípade predpokladaných kumulatívnych a synergických účinkov. Niektoré z nich je nutné tiež presnejšie kvantifikovať (napr. meranie úrovne hluku atď.). |
||||
|
Poďakovanie |
||||
|
Ďakujem za pripomienky, cenné rady a materiál RNDr. Mirkovi Bohušovi, PhD., za mnohé terénne údaje ornitológom Soni Nuhlíčkovej, Štefanovi Benkovi a Andrejovi Chudému, Trenčianskemu múzeu a za rady ohľadom štatistického spracovania RNDr. Petrovi Degmovi, CSc. |
||||
|
References |
||||
|
Begon M, Harper LJ
& Townsend RC, 1996: Ecology. Blackwell science, Oxford, 3. vydanie, 1068 pp.
Danko Š, Darolová A & Krištín A, 2002: Rozšírenie vtákov na Slovensku. Veda, Bratislava, 686 pp. Deván P & Májsky J, 1985: Ochrana prírody v okrese Trenčín. Vydavateľstvo Obzor, Bratislava, 112 pp. Gregorová G & Filová V, 2004: Štatistické metódy v geografii. Geografika, Bratislava, 117 pp. Hudec K & Černý W (eds), 1977: Fauna ČSSR. Ptáci II. Academia, Praha, 866 pp. Hudec K (ed.), 1983: Fauna ČSSR. Ptáci 3/1, 3/2. Academia, Praha, 1 236 pp. Hudec K (ed.), 1994: Fauna ČSFR. Ptáci 1. Academia, Praha, 671 pp. Jambor R, 2003: Vtáky trenčianskeho úseku Váhu. Chránené územia Slovenska, 57: 28–31. Janda J & Řepa P, 1986: Metody kvantitativního výskumu v ornitologii. SZN, Praha, 157 pp. Odum PE, 1977: Základy ekologie. Academia, 3. vydanie, Praha, 733 pp. Pelikán J, 1993: Přehled obecné ekologie. Ediční středisko VŠVF, Brno, 153 pp. Voous KH, 1973: List of recent Holarctic bird species. Non – Passerines. Ibis, 115: 612–638. Voous KH, 1977: List of recent Holarctic bird species. Passerines. Ibis, 119: 223–250, 376-406. |
||||
|
|