Folia faunistica Slovaca 20 (2) 2015: 113–129
Úvod
Prvé pokusy o klasifikáciu typov tokov na základe spoločenstiev pošvatiek sa objavili v prácach Berga (1948) a Illiesa (1961). Illies & Botosaneanu (1963) a Hawkes (1975) následne vypracovali klasifikáciu riečnej zonácie. Vplyvom rozmanitých environmentálnych faktorov ovplvňujúcich zonáciu pošvatiek sa zaoberali Illies (1952), Levadinov (1981), Braukman (1987), Statzner (1987), Krno (1984), Soldán et al. (1998) a Krno & Holubec (2009).
Existujú dva základné prístupy ku klasifikácii tokov – zonačný (v prácach vyššie uvedených autorov) a kontinuálny Maitland (1966) a Vannote et al. (1980), ktorí sa ako prvý zaoberali teóriou riečneho kontinua, ktoré charakterizujú plynulou zmenou fyzikálnych podmienok a následnou sériou odpovedí v štruktúre biocenóz , od prameňa k ústiu. Vannote et al. (1980) charakterizovali tečúce vody ako starobylý, ale flexibilný systém, ktorý rýchlo odráža zmeny prostredia, ich dlhá brehová línia podmieňuje existenciu otvoreného vodného ekosystému, úzko súvisí s suchozemskými ekosystémami. Túto teóriu vo svojich prácach potvrdzujú Naimann et al. (1987), Minshall et al. (1992), Krno et al. (1996). Rozdiel zonálneho a kontinuálneho pohľadu na hodnotenie vodných biotopov prekonávajú vo svojej práci Perry & Schaffer (1987), ktorý tvrdia, že gradient zmien taxonomického zloženia makrozoobentosu v tokoch je postupný, avšak spojený s náhlymi zmenami, ktoré sú sprevádzané zjavným zvýšením diverzity. Ku kombinácii oboch pohľadov sa prikláňajú aj Hildrew & Townsend (1987).
Výborným indikátorom zonácie, zdravia horských a podhorských tokov a prírodných podmienok ovplyvňujúcich vodné ekosystémy sú pošvatky. Pošvatky sú užitočné biologické indikátory pôvodnosti a kvality riečnej krajiny, odrážajúce degradáciu toku a využitie krajiny (Krno, 2009; Krno, Žiak, 2012). K spoznaniu súčasného stavu spoločenstiev pošvatiek horských a podhorských tokov Nízkych Tatier prispel predovšetkým Krno (1978, 1982, 1987).
Erózia pôdy je dôsledkom geomorfologických vlastností povodia (Štefanovič et al. 2007). Erózia pôdy je prirodzený jav, pod vplyvom veľkého množstva faktorov (geomorfologické, geologické, pedologické, meteorologické, hydrologické, biologické, atď.). Okrem prirodzeného aspektu zohráva veľmi negativnu úlohu antropogénny faktor. Erózia sa prejavuje buď priamym účinok v oblasti erózie, ktorá sa prejavuje degradáciou pôdy a prírodného prostredia, alebo nepriamym vplyvom mimo zónu erózie, čo sa prejavuje transportom sedimentov prostredníctvom hydrografickej siete povodia (Štefanovič et al. 2007). Okrem ekonomickej činnosti člověka ovplyvňujú eróziu pôdy aj niektoré neekonomické činnosti – rekreácia, najmä masová výstavba lyžiarskych stredísk atď. V dôsledku toho sú hospodárske a sociálne dôsledky erózie pôdy veľmi významné. V súvislosti s kandidatúrou Tatier na ZOH sa dostávala do popredia otázka vplyvu výstavby lyžiarskych a ubytovacích zariadení na okolité ekosystémy. Rozsiahly výskum daného problému robil v Alpách Frantz (1979). Cernusca (1987) sa zaoberal vplyvom lyžiarskych zariadení v Alpách na suchozemskú flóru a faunu. V Tatrách sa vplyvom zjazdoviek na vegetáciu zaoberali botanici Čunderlíková & Marhold (1984). Kocian (1992) sledoval negatívny vplyv zjazdového lyžovania na výskyt suchozemských stavovcov. Midriak (1993) sledoval v Nízkych Tatrách zvýšený odnos pôdy vplyvom odlesnenia. Negatívny vplyv zjazdoviek na krajinu priťahuje odborníkov z viacerých hľadísk. Krajinno-ekologické predpoklady pre návrh lyžiarskych tratí v oblasti Nízky ch Tatier spracovala Hrnčiarová (1996).
Vplyvom odstraňovania lesnej vegetácie a následnej erózie na toky a ich oživenia sa zaoberali Gollday et al. (1992). Poukazujú predovšetkým na zvýšený stupeň mineralizácie a tým spôsobenú zmenu chemizmu vody vplyvom erózie. Minshall et al. (1992) a Krno et al. (1995), Krno & Žiak (2012) sa zaoberajú vplyvom erózie na vodné spoločenstvá. Ako hlavný vplyv na spoločenstvá makrozoobentosu uvádzajú obrusovanie nárastov a upchávanie intersticiálnych priestorov a tým zmenu v potravných gildách spoločenstiev.
Cieľom tejto práce bolo zachytiť zmeny v druhovom zložení taxocenóz pošvatiek, ekologických metrík meniacich sa v rámci ekologického gradientu (nadmorská výška, vodnatosť) od vysokopoložených horských bystrín až po ústie podhorského toku Demänovky do Váhu (Nízke Tatry). Zároveň sme sa snažili zachytiť vplyv zvýšenej erózie na spoločenstvom pošvatiek. Zvýšenú eróziu sme predovšetkým sledovali na prítoku Demänovky (Priečne) v rokoch 1995–1997, ktorý zberá vody z lyžiarsky nadmerne využívaných svahov Chopka.
Materiál a metódy
Charakteristika územia
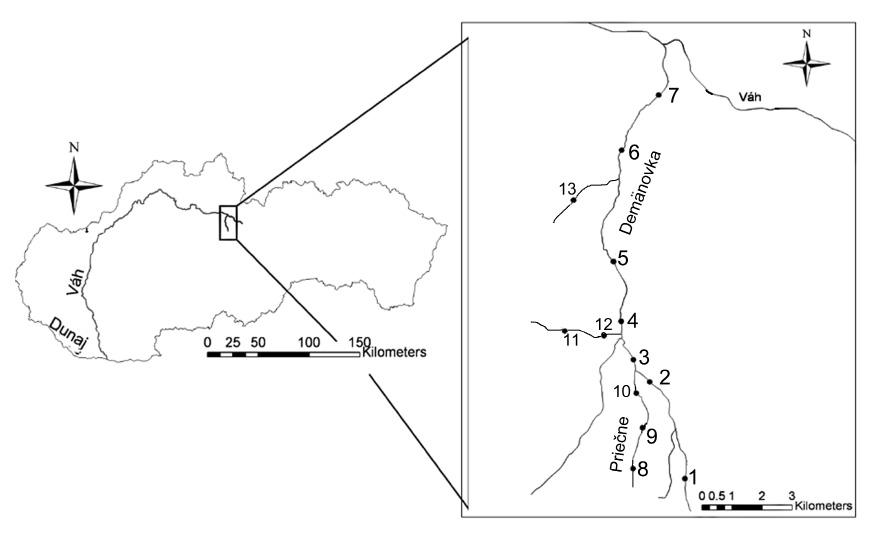
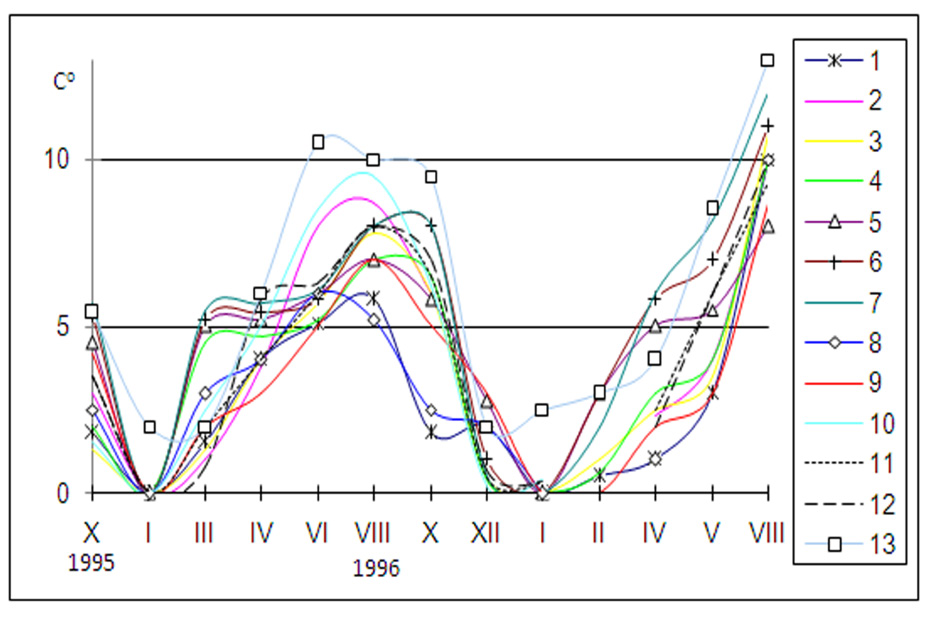
Riečka Demänovka (lok. 1–7) je ľavostranným prítokom horného Váhu (Obr. 1). Preteká cez Demänovskú dolinu, ktorá vybieha na sever z hlavného hrebeňa Nízkych Tatier. Pramenná oblasť najvyššie položených prítokov leží v priestore od Poľany (1890 m) až po Krupovú hoľu (1927 m). V severných dolinách skupiny Ďumbier, bolo v poslednej dobe viac ľadovcov (Droppa 1972). Pod Lúčkami sa Demänovská dolina výrazne zužuje a prehlbuje do bralnatého kaňonu, pričom horniny kryštalinika prechádzajú do sedimentárnych súvrství križňanského príkrovu (Lacika 1992). Na tomto rozhraní sa nachádza veľké množstvo ponorov Droppa (1957). Vďaka vode, ktorá sa tratí v ponoroch, vznikol unikátny, 22 km dlhý, jaskynný systém s 9 horizontálnymi úrovňami. Ako riečka Demänovka opúšťa Demänovskú dolinu, vstupuje do flyšovej Liptovskej kotliny, kde vytvára typické riečne terasy (Droppa 1970). Povodie riečky Demänovky leží na rozhraní chladnej a mierne teplej klimatickej oblasti (Jakál & Mazúr 1980). Riečka Demänovka je tokom 4. rádu. Od prameňa až po ústie do Váhu prekonáva vzdialenosť 19 km a približný výškový rozdiel je 1000 m. Plocha povodia, ktorá dosahuje 61 km2, má pretiahnutý tvar, pomer jej šírky k dĺžke toku je 1:3. Lesnatosť tejto oblasti dosahuje 60 %, pričom dominantnou drevinou je smrek obyčajný. V povodí Demänovky spadne priemerne 1072 mm zrážok. Priemerné hodnoty špecifického odtoku dosahujú 18 l.s–1 na km2 (Balco 1977). V roku 1996 bol priemerný prietok 1,218 m3.s–1, pričom jeho hodnoty sa pohybovali od 0,11 m3.s–1 do 11,67 m3.s–1. V roku 1997 bol priemerný prietok 0,816 m3.s–1, pričom jeho hodnoty sa pohybovali od 0,17 m3.s–1 do 12,61 m3.s–1. Maximálny prietok býva v apríli a minimálny v zimnom období. Na základe uvedených charakteristík môžeme povodie riečky Demänovky zaradiť k stredohorskému typu B1 (Šimo 1972). Stredohorský charakter Demänovky podmieňujú tiež jej špecifické teplotné pomery. Riečka prijíma početné prítoky, pričom väčšina z nich pramení vo vysokých polohách. Tečú pritom úzkymi zalesnenými údoliami. Ich teplota okrem letných mesiacov zvyčajne nepresahuje 8 °C. Na základe teplotného režimu tokov (Obr. 2) môžeme pomerne dobre rozlíšiť chladnú periódu (december až marec) a teplú periódu (máj až október). Maximálna letná teplota v horských bystrinách sa pohybovala okolo 10 °C. Letné maximum v podhorských tokoch dosahovalo len 13 °C. Nízke teploty vody v povodí Demänovky sú ovplyvnené orientáciou Demänovskej doliny na sever a podzemnými vodami vyvierajúcimi z jaskynných systémov.
Charakteristika základných environmentálnych dát tokov je uvedená v tabuľke 1. Potok Priečne (lok. 8–10), najmä jeho dolná časť, pretekal cez lyžiarsky terén, priteká spod Chopka. Potok Radová (11, 12) je ľavostranný kaňonovitý prítok spod Sinej. Rakový potok (13) preteká cez PR Jelšie.
Metodika
Počas dvoch rokov terénneho výskumu (október 1995 – august 1997) sme v povodí Demänovky sledovali 13 vybraných stacionárov.
Kvalitatívne odbery makrozoobentosu sme robili na 13 stanovištiach v pravidelných štvrťročných intervaloch. Pri odberoch sa použila tzv. „kicking technique“, ktorú prvý krát použil Hynes (1961). Vzorky sme odoberali naprieč celým tokom.
Kvantitatívne odbery makrozoobentosu a detritu sme robili počas dvoch rokov v dvojmesačných intervaloch na lokalitách 2 a 10. Na potoku Priečne (lokalita 10) sme sledovali vplyv erózie na štruktúru spoločenstva makrozoobentosu. Lokalita 2 (Demänovka) nebola vystavená eróznemu tlaku a slúžila ako kontrola. Na kvantitatívne odbery sme použili Kubíčkov bentometer trojuholníkového tvaru, o ploche 0,1 m2 (Helan et al. 1973). Z hrubého substrátu scaly (makro- a mezolital) sme odobrali vzorku z plochy 0,3 m2. Kvantitatívny odber z jemného substrátu (akal, piesok) bol odobraný z plochy 0,1 až 0,2 m2.
Na lokalitách 2 a 10 sme zároveň s kvantitatívnymi vzorkami makrozoobentosu odoberali aj vzorky hrubého bentického material (CBOM). Vzorky jemného (FBOM) a ultrajemného (UFBOM) materiálu sme odberali pomocou trubice zarazenej do dna, o priemere 19 cm (na substráte skaly 3 ×, na substráte piesok 1–2 × (Krno et al. 1996). Partikulárny organický material (POM) bol vysušený 3,5 hod pri 105 °C, potom bol na porcelánových miskách žíhaný v muflovej peci 3,5 hod pri 550 °C a odvážený ako AFDM. Pri jarných, letných a jesenných odberoch sa na týchto stanovištiach brali vzorky vody na stanovenie doplnkových fyzikálno-chemických parametrov (pH, vápnika, dusičnanov a BSK5).
Pri vyhodnotení štruktúry spoločenstiev pošvatiek sme použili nasledovné metriky:
Počet druhov (S), α-diverzitu, saprobný index (Si) – Krno (2013), index druhovej diversity (H) – (Shannon & Weaver 1949), index vyrovnanosti (e) Pielou (1966), priemerné skóre pošvatiek (SAS), a celkové skóre pošvatiek (STS) boli vyhodnotené podľa Krna (2007), LN_index (erózny index) – vyhodnotený podľa Krna et al. (2015).
Vyššie uvedené ekologické metriky sme stanovili pre jednotlivé sezónne odbery a výslednú hodnotu na základe celoročných údajov.
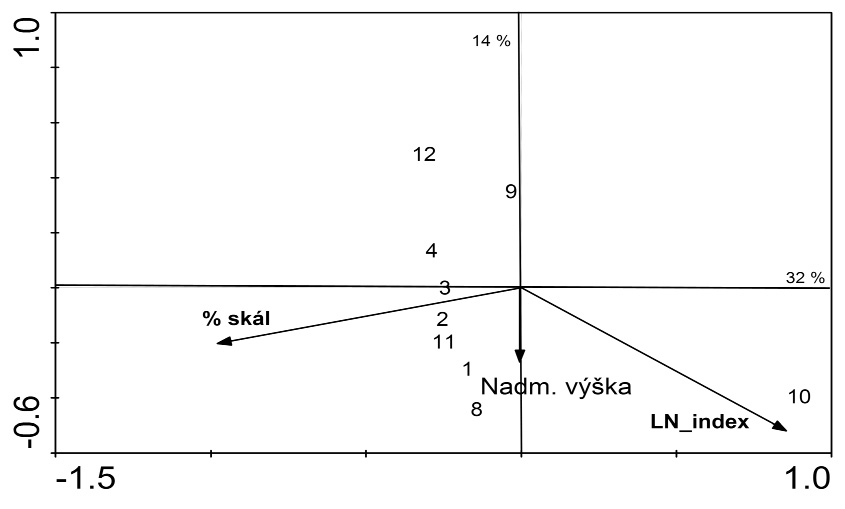
Podobnosť spoločenstiev pošvatiek na základe relatívnej druhovej početnosti sme použili PCA, Na analýzu vzťahov pošvatiek povodia Demänovky a vybraných environmentálnych faktorov prostredia sme sa rozhodli použiť kanonickú korešpondenčnú analýzu (CCA). Na selektovanie najvýznamnejších faktorov sme použili metódu „forward selection“. Každú vybranú premennú sme testovali Monte Carlo permutačným testom s 999 permutáciami a zistili sme, či jej vplyv na rozmiestnenie druhov v ordinačnom priestore je náhodný alebo štatisticky významný (Ter Braak 1991).
Výsledky
Taxocenózy pošvatiek a environmentálne premenné
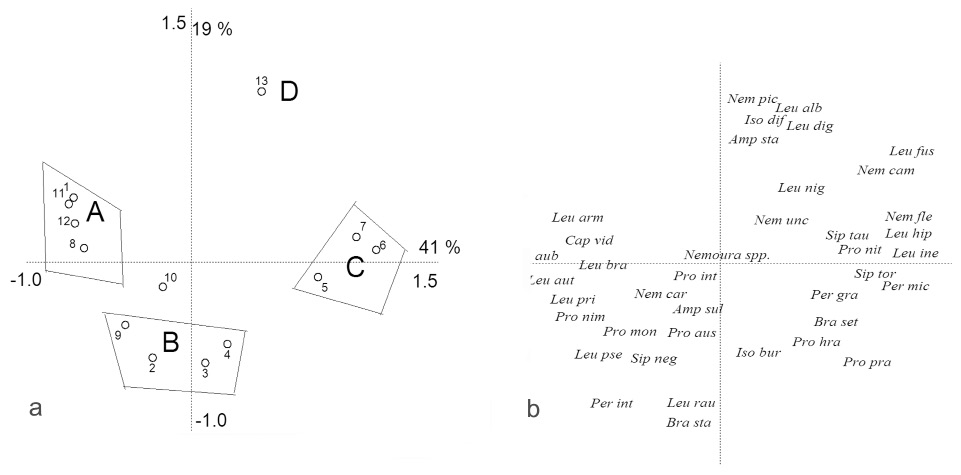
Výskum spoločenstva pošvatiek sme robili na 13 lokalitách povodia Demänovky (Tab. 2). V období výskumu sme z odobraného bentosu determinovali 47 taxónov pošvatiek patriacich do 11 rodov. Najpočetnejšie boli zastúpené rody Leuctra a Protonemura. Na základe výsledkov predchádzajúcich PCA analýzy (Obr. 3a, b) môžeme vyčleniť 4 základné spoločenstvá pošvatiek patriace do dvoch hlavných skupín:
I. Spoločenstvá pošvatiek horských tokov:
A. Spoločenstvá pošvatiek vysokopoložených horských tokov a teplotne inverzných tokov kaňonovitých dolín.
Ide o spoločenstvá lokalít 1, 8 na silikátovom podloží (nad 1300 m n. m., Demänovka, p. Priečne) a lokalít 11, 12 (ležiace krasovom kaňone nad 800 m n. m., Radový p.), ktorých je substrát tvorený prevažne balvanmi a skalami, s vysokou pokryvnosťou machov (hlavne Palustriella commutata).
Charakteristické sú relatívne vysokými indexmi diversity, SAS a veľkou vyrovnanosťou druhového zloženia a nízkym sapróbnym indexom (Si) (Tab. 3). Charakteristickým dominatným druhom týchto horských bystrín je Leuctra pusilla a typickým Protonemura brevistyla, P. auberti.
Typickými druhmi silikátových tokov sú rod Brachyptera a druhy Protonemura nimborum, Leutra armata, Capnia vidua a Perlodes intricatus. Pre karbonátové toky sú Leuctra braueri, L. prima a Diura bicaudata.
B. Spoločenstvá pošvatiek pôvodných vodnatejších horských tokov.
Sú to spoločenstvá charakteristické pre lokality 2, 3, 4 (Demänovka), 9, kedysi sem patrila aj lokalita 10 (Priečne) (800 – 1000 m n.m.). Substrát je tvorený balvamni a skalami s menšou pokryvnosťou machov (Brachythecium rivulare) ako u predchadzájúceho typu habitatu. Oproti predchadzajúcemu spoločenstvu sa vyznačuje o niečo nižšími ekologickými indexami s výnimkou Si (Tab. 3). Typickými druhmi sú Brachyptera starmachi, Protonemura nimborum, Leuctra rauscheri, Perlodes intricatus a Siphonoperla neglecta.
II. Spoločenstvá pošvatiek podhorských tokov.
C. Spoločenstvá pošvatiek podhorských vodnatejších tokov.
Na riečke Demänovke majú charakter podhorských tokov (600 – 800 m n.m.) lokality 5, 6 a 7. Substrát tvoria balvany a skaly s prevahou skál, s prechodom až do hrubého štrku, pričom je tu minimálny výskyt machu. Charakteristické sú nízkymi ekologickými indexami, najmä mierne eutrofizovaný dolný tok rieky – polia, a zvýšeným saprobným indexom. Pre spoločenstvá sú vlastné druhy Nemoura flexulosa, Perlodes microcephalus, Perla grandis, Siphonoperla torrentium, pričom extrémne dominoval druh Leuctra inermis.
D. Spoločenstvo pošvatiek podhorského pramenného toku (Rakový p.) s piesočnato-bahnitým dnom, pretekajúci pretekajúci jelšovím lesom.
Takýto typ spoločenstva je lokalizovaný na stanovišti 13. Lokalita je charakteristická nízkym indexom diverzity i vyrovnanosti. Vyskytovalo sa na nej najmenej druhov zo všetkých lokalít (13). Dominujú tu predovšetkým dva druhy Amphinemura standfussi a Nemurella pictetii. Ako menej významné ich dopĺňajú druhy Leuctra albida, L.digitata L.fusca, L. nigra a Isoperla difformis.
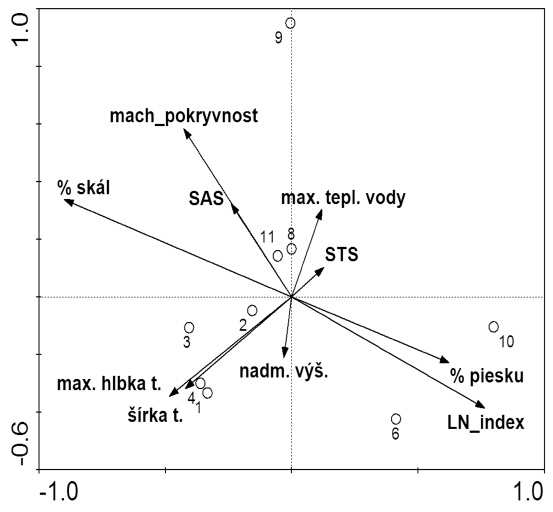
PCA analýza premených je zachytená na Obr. 4, kde koreluje pokryvnosť machu s pôvodnosťou toku (SAS), erózny index (LN) s podielom jemného substrátu, a samozrejme hĺbka so šírkou toku.
Kanonickú korešpondenčnú analýzu (CCA) sme použili na analýzu vzťahov pošvatiek povodia Demänovky a vybraných environmentálnych faktorov prostredia (Obr. 5). Použili sme kvantitatívnu druhovú maticu a maticu hodnôt 12 environmentálnych premenných (nadmorská výška, spád, šírka, hĺbka toku, pokryvnosť machov, percentuálne zastúpenie substrátu balvany a skaly resp. piesku, maximálna teplota a niektoré ekologické metriky). Kumulatívna percentuálna variancia druhových dát na dvoch osiach bola 46 %. Najvýznamnejšie enviromentálne premenné, ktoré ovplyvňujú ekologické postavenie úsekov riečky Demänovky a jej prítokov sú nadmorská výška (tá koreluje so spádom toku, teplotným režimom), podiel hrubého substrátu (mierne koreluje s pokryvnosťou machov) na dne a LN index, ktorý súvisí s eróziou a podielom jemného substrátu.
Antropogénne vlyvy na taxocenózy a ekologické metriky pošvatiek
Vplyv človeka sa premietol aj do spoločenstiev pošvatiek riečky Demänovka (lok. 1–7). Prejavil sa popri prirodzenej zvýšenej eutrofizáciii v rámci riečneho kontinua aj kontinuálnym či bodovým organickým znečistením najmä v dolnom toku rieky. Tieto závery jednoznačne potvrdzujú ekologické metriky (Tab. 4), kedy sme v dolnom toku zaznamenali diskontinuitné zhoršenie všetkých metrík, najmä však sapróbneho indexu, indexu diverzity a celkovej biodiverzity pošvatiek.
Vo výskumnom období 1995–1997 sme sa predovšetkým zamerali na podrobnejšiu analýzu dvoch profilov ležiacich v pôvodnom povodí Demänovky (lok. 2) a povodí potoka Priečne, narušenom eróziou (lyžiarske terény, hotelová výstavba a cestné komunikácie).
Demänovka na Lúčkach (lokalita 2)
Fyzikálno-chemické (Tab. 4) parametre boli robené v mesiacoch máj a júl, október 1996 a 1997. Koncentrácia vodíkových iónov kolísala od 5,7 do 7,1. Obsah vápnika sa pohyboval od 4 do 9 mg/l. Množstvo dusičnanov vo vode sa pohybovalo od 1,7 do 2,4 mg/l. Hodnoty BSK5 kolísali okolo 1 až 1,5 mgO2/l.
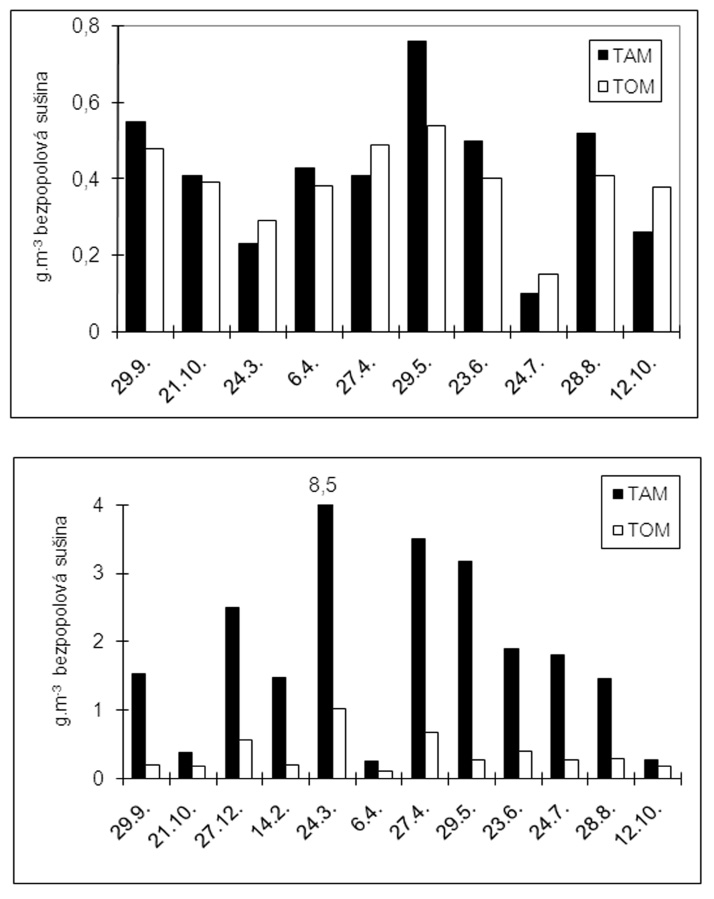
V tomto období sme sledovali aj sezónnu dynamiku transportovaného organického materiál (TOM) a transportovaného anorganického materiálu (TAM) (Obr. 6a). Namerané množstvá transportovaného materiálu dosahovali maximá na jar a začiatkom jesene. Pomer transportovanej organickej a anorganickej zložky bol približne vyrovnaný s miernou prevahou organickej zložky. Výnimkou bola jar 1997, kedy množstvo TAM takmer dvojnásobne prevyšovalo množstvo TOM. Množstvá TAM a TOM sa pohybovali od 0,10 g/m3 v zime po 0,76 g/m3 pri jarnom maxime.
Na mikrohabitatoch makro-mezolital bolo množstvo CBOM 3× nižšie (priemer 11,4 g.m–2) ako na substrátoch akal-psamal (34,2 g.m–2). Najväčšie množstvá detritu sú na substráte scaly, namerali sme ho neskoro na jar a začiatkom leta. Na celkovom množstve bentickej organickej hmoty sa podieľali krytosemenné rastliny v priemere 74,6 %, ihličie 11,1 %, drevo 13,7 %. CBOM na mikrohabitatoch akal-psamal bola najvyššie na jar a na jeseň. Výnimkou boli vysoké množstvá namerané v lete 1997, po povodniach. Substrát akal-psamál zachytil päťnásobné množstvo (480,4 g.m–2) jemného organického materiálu (FBOM+UFBOM) v porovnaní so substrátom makrolotál-mezolital (85,1 g.m–2) (Gašperčíková 1998).
Makrozoobentos v makro-mezolitále dosahoval priemernú denzitu 1621 ex.m–2. Plecoptera bola reprezentované 12% (194 ex.m–2). Na mezohabitate akal-psamal bola priemerná abundancia makrozoobentosu 1617 ex.m–2. Plecoptera sa podieľali len 5% (81 ex.m–2). Spoločenstvo pošvatiek sa neodlišovalo od ostatných horských bystrín.
Potok Priečne na Lúčkach (lokalita 10)
Reakcia vody ukazuje na mierny posun hodnôt pH smerom do zásaditej oblasti (Tab.4). Namerané hodnoty pH v roku 1996 boli 6,5 v máji a 7,3 v októbri, v roku 1997 boli 7,3 v máji a 7,5 v októbri. V roku 1997 sa oproti roku 1996 mierne zvýšil obsah vápnika z 8 mg/l na jar 1996 na 12 mg/l na jar 1997 a výrazne, rádovo o jedno desatinné miesto, poklesol obsah amoniaku a amónnych solí. Obsah dusičnanov vo vode kolísal okolo priemernej hodnoty 3.0 mg/l.
Množstvo transportovaného anorganického materiálu (TAM) je na erodovanom toku niekoľkonásobne vyššie ako množstvo transportovaného organického materiálu (TOM). Najvyššie hodnoty TAM a TOM boli na jar. Namerané hodnoty TAM sa pohybovali od 0,26 g/m2 do 10,70 g/m2. Namerané hodnoty TOM sa pohybovali rádovo od 0,11 g/m2 do 0,68 g/m2 (Obr. 6b).
Celkové množstvo bentického detritu na tomto úseku v porovnaní z Demänovkou bolo 1,5 x nižšie. Priemerné množstvo CBOM na makro-mezolitale bolo 5,8 g/m2 a na jemnejších substrátoch až 28,2 g/m2. Z hľadiska sezónnej dynamiky bolo najviac organickej hmoty usádzanej na jar a na jeseň. Na celkovom množstve bentickej organickej hmoty sa podieľali krytosemené rastliny v priemere 59,8 %, ihličie 16,2 %, drevo 24 %. Sezónne maximá sme namerali najvyššie na jar a na jeseň. Najnižšie hodnoty boli v lete. Substrát akal-psamál zachytil 15-násobné množstvo (410,5 g.m–2) jemného organického materialu (FBOM+UFBOM) v porovnaní so substrátom makrolotál-mezolital (25,1 g.m–2) (Gašperčíková 1998).
Priemerná abundancia makrozoobentosu zistená na mezohabitate macro-mesolithal bola 1968 ex/m2. Početne zastúpené boli aj Plecoptera v priemere 277 ex/m2. Ich podiel na množstve bentických organizmov bol 14 %.
Na substráte piesok bola priemerná abundancia makrozoobentosu len 608 ex/m2. Dominantne boli zastúpené Plecoptera 20% s priemernou abundanciou 119 ex/m2.
Počas výskumu sme z odberov na lokalite 10 determinovali 18 taxónov z radu Plecoptera. Priemerná abundancia jedincov bola v danom období 198 ex/m2. Najpočetnejšie zastúpenie spomedzi pošvatiek mali taxóny Nemoura spp. a Leuctra rauscheri (Tab. 2), ktoré boli na danej lokalite eudominantné. Dominantne zastúpené boli Protonemura auberti 11,5% a Protonemura austriaca 12,7%. Žiaden zo zvyšných druhov pošvatiek neprekročil 10% podiel). Podiel rodov Nemoura a Leuctra bol neporovnateľne vyšší, než na ostatných lokalitách, čo sa premietalo do vyššieho erózneho LN_indexu (Tab. 4), podobné zvýšené hodnoty sme zaznamenali v dolnom toku Demänovky, pretekajúci cez poľnohospodársku krajinu.
Diskusia
Vo výskumnom období október 1995 – august 1997 sme z 13 lokalít povodia Demänovky determinovali 47 taxónov pošvatiek.
Z povodia Ľupčianky, ktoré sa nachádza rovnako ako povodie Demänovky v Nízkych Tatrách, determinoval Krno (1982) spolu 47 druhov pošvatiek (Tab. 5). Autor uvádza ako dominantné druhy Protonemura nimborum, P.montana, P.praecox, P.hrabei, Leuctra autumnalis, L.inermis, L.albida, L.fusca, L.rauscheri, Isoperla sudetica, I.buresi a Dinocras cephalotes. Z druhov determinovaných z povodia Ľupčianky, v povodí Demänovky absentovali druhy Taeniopteryx auberti, Nemoura monticola, Leuctra aurita, L.moselyi, Perla marginata a Dinocras cephalotes.
Na rieke Belej a jej významnejších prítokoch, ktoré zberajú vody z južných svahov Západných a Východných Tatier, zistil Krno (1984) počas 5-ročného výskumu spolu 59 druhov pošvatiek (Tab. 5). V povodí rieky Belej dominovali druhy Brachyptera starmachi, B. seticornis, Rhabdiopteryx neglecta, Protonemura auberti, P.autumnalis, P.brevistyla, P.intricata, P.montana, P.nimborum, P.praecox, Amphinemura standfussi, A.sulcicollis, Leuctra aurita, L.armata, L.autumnalis, L.handlirschi, L.nigra, L.rauscheri, L.rosinae, Isoperla sudetica, I.oxylepis a Arcynopteryx dichroa. Z daných dominantných druhov sme na povodí Demänovky nezachytil druhy Leuctra rosinae a Rhabdiopteryx neglecta. Z ostatných druhov povodia Belej som v povodí Demänovky nezaznamenal Taeniopteryx auberti, Leuctra aurita, L.major, L.moselyi, Arcynopteryx dichroa, Isoperla rivulorum, Chloroperla kisi a Dinocras cephalotes.
V povodí Demänovky sme zachytili 3 druhy, ktoré sa v povodiach Ľupčianky a Belej nevyskytovali: Nemoura carpatica, Isoperla difformis a Siphonoperla taurica.
Na 127 lokalitách povodia horného Váhu, zaznamenal Krno (1987) počas 10 ročného výskumu 61 druhov radu Plecoptera.
V porovnaní s povodiami prítokov horného Váhu (Belá, Ľupčianka, Revúca (Krno 1978)), je v povodí Demänovky zaujímavá práve absencia druhu Dinocras cephalotes. Ak sú vajíčka tohto druhu vystavované teplote pod 4°C upadajú do stavu dormancie. Na prekonanie stavu dormancie a vyliahnutie larvy je potrebná teplota minimálne 10 °C (Zwick 1996a). Pri sledovaní nemeckej populácie druhu Dinocras cephalotes zistil Zwick (1996b) viac ako 90% úspešnosť liahnutia vajíčok v teplotnom rozmedzí 12 – 18°C. Absencia tohto druhu v povodí Demänovky pravdepodobne súvisí s nízkymi maximálnymi teplotami v lete (TMAX 1996 8,6 °C, TMAX 1997 9,6 °C). Hydrotermický režim povodia Demänovky obmedzuje výskyt aj ďalších teplostenotermných a prevažne letných druhov ako napr. Perla marginata, P.burmeisteriana, Protonemura autumnalis, Leuctra albida, L.aurita, L.hippopus, L.mortoni, L.moselyi a Isoperla oxylepis. Tento spôsob eliminácie teplomilnejších druhov v izolovanom povodí Demänovky naznačuje analógiu s mechanizmom vytláčania pôvodných teplomilnejších druhov z povodí západokarpatskej sústavy v chladnejších geologických obdobiach. Čeľaď Perlidae, do ktorej druh Dinocras cephalotes patrí, dosahuje najvyššiu diverzitu v teplých až tropických podmienkach (Zwick 1996b). Jediným zástupcom čeľade na Demänovke bol chladnomilný druh Perla grandis. Krno (1982, 1984) ju uvádza v povodí Ľupčianky do 900 m n.m., v povodí Belej do 1300 m n.m. Auberta (1959) ju zo švajčiarskych Álp uvádza až do 2000 m n. m.. Na Demänovke sa vyskytovala na podhorskom úseku toku pod Demänovskou ľadovou jaskyňou (740 m n. m.). Nad týmto úsekom vyvierajú do riečky Demänovky krasové podzemné vody. Podľa Zaťka (1980) môže podzemná voda výrazne ovplyvňovať teplotu povrchových vôd, zmierňovaním denných, respektíve ročných výkyvov. Tento vplyv môže byť tak výrazný, že spôsobuje inverziu teploty vody v letných mesiacoch. V závislosti od režimu toku môže mať Perla grandis 2 až 5 ročný vývinový cyklus. Na hornom toku rieky Necker vo švajčiarskych Alpách má Perla grandis 4 ročný vývinový cyklus (Frutiger & Imhof 1997). Stacionár pod Demänovskou ľadovou jaskyňou má podobný teplotný režim.
Významným ľavostranným prítokom Váhu, riekou Turiec, ktorým sa zaoberal Krno et al. (1996). Z tokov povodia Turca, ktoré zbierajú vody z Turčianskej kotliny a priľahlých pohorí – Veľkej Fatry, Kremnických vrchov, Malej Fatry a pohoria Žiar, uvádza autor až 64 druhov pošvatiek, čo predstavuje 2/3 slovenskej plekopterofauny. Táto druhová pestrosť je pripisovaná rôznorodosti biotopov. V epiritráli Demänovky sa na rozdiel od epiritrálu Turca nevyskytovali druhy Taeniopteryx nebulosa, Nemoura monticola a Arcynopteryx dichroa. Metaritrál Turca bol bohatší o druhy Brachyptera risi, Taeniopteryx auberti, Nemoura babiagorensis, N.cinerea, N.marginata, Leuctra moselyi, Capnia bifrons, Perla marginata a Dinocras cephalotes. V povodí Demänovky nie je zóna hyporitrálu
V povodí Demänovky sme zachytili západokarpatské endemity Brachyptera starmachi a Leuctra pusilla a karpatský endemit Nemoura carpathica. Zaujímavý bol výskyt druhu Siphonoperla taurica, ktorý je doposiaľ známy len z pohorí, ktoré majú bezprostredný kontakt s Podunajskou nížinou (Krno et al. 1996).
Dnešné rozšírenie pošvatiek je výsledkom dlhého historického vývoja, pri ktorom prešiel druh v úzkom vzťahu s meniacim sa geografickým územím. Obzvlášť paleopotamologické, paleogeomorfologické a paleoklimatologické pomery určujú dnešný stav rozšírenia asociácíí Plecoptera (Krno et al. 1996).
Z povodia Demänovky sme na základe PCA, CCA analýzy vyčlenili 4 základné spoločenstvá pošvatiek patriace do dvoch hlavných skupín: spoločenstvá horských tokov a spoločenstvá podhorských tokov. Podobné taxocenózy potočníkov viazané na horské a podhorské úseky toku Demänovky popísali Beracko et al.. (2012).
Prác, ktoré sa zaoberajú spoločenstvami jednotlivých typov tokov je už veľké množstvo. Už Brinck (1949) presne definoval spoločenstvá pošvatiek v rôznych typoch tokov. Podrobne popisuje v akom úseku aký druh dominuje. Avšak toky zaraďuje do empirických skupín ako lesný tok, meandrujúci tok a iné. Larvy pošvatiek boli typické pre horné lesné úseky škandinávskych tokov..
Raušer (1964) sa bližšie zaoberal asociáciami pošvatiek bývalého Československa. Areál bývalej ČSSR rozdelil horizontálne aj vertikálne na 3 základné typy povodí a početné varianty, ktoré sa navzájom líšia diferenciačnými druhmi. Autor rozlišuje lenitické povodia, vody pahorkatín a predhorí a vody hôr. Na území Karpát rozlišuje dve varianty horských vôd: vysokohorská varianta (Vysoké Tatry) a horská karpatská varianta.
Illies (1961), Illies & Botosaneanu (1963) zhrnuli znalosti o biocenózach a fyzikálno-chemických podmienkach lotického systému do hydrobiologickej typológie tokov. Bez ďalšieho členenia ritrálu, uvádzajú ako charakteristické čeľade pošvatiek tohto pásma Capnidae, Leuctridae, Nemouridae, pre pásmo potamálu predovšetkým čeľade Perlodidae a Perlidae.
Prístup autorov ku klasifikácii tokov je až do 70. rokov minulého storočia dosť individuálny. Mnoho údajov o tokoch nachádzame v systematických prácach, v ktorých autori väčšinou uvádzajú kde a v akom type toku získali imága a larvy toho ktorého druhu. Hydrobiologickú typológiu tokov akceptujú až neskôr publikované práce.
Hawkes (1975) vypracoval klasifikáciu riečnej zonácie, pričom nadviazal na štúdie longitudinálnej zonácie založené na ichtyofaune. Práca vychádzala predovšetkým z fyzikálnych (spád toku, teplota, substrát, veľkosť toku) a chemických (rozpustený kyslík, rozpustené nutrienty, tvrdosť vody) charakteristík.
S novým prístupom k typológii tokov na tzv. choriotopy prišiel Braukman (1987). V jeho indikačnom systéme tvoria významné postavenie larvy pošvatiek. Diferencuje druhy pošvatiek indikujúce typ podložia toku. Krno (1992) a Helešic (1995) uvádzajú naopak, že pošvatky sú málo citlivé na charakter podložia pôvodných povodí. Avšak súhrné spracovanie rozšírenia pošvatiek Slovenska (Žiak, 2012) signalizuje, že tomu tak celkom nie je a zaznamenáva isté rozdielnosti v dominancii rôznych druhov pošvatiek v povodiach s rozličným podložím (napr, jadrové, sopečné, vápencové alebo flyšové). V povodí Demänovky sme zaznamenali výrazné zmeny v štruktŕe karbonátových tokov v porovnaní s jadrovými, viaceré údaje napr. pri druhoch Leuctra braueri, L. prima a Capnia vidua odpovedajú pozorovaniam autorov Graf et al. (2009). Pri rode Brachyptera podobne ako v tatranských tokoch (Krno et al. 2015) sme však zaznamenali, že tento rod sa vyslovene vyhýba vápencovým tokom. Predátori rodu Diura uprednosňovali skôr karbonátové toky na rozdiel od predátora Perlodes intricatus.
U nás sú relatívne dobre opísané spoločenstvá pošvatiek predovšetkým horských a podhorských tokov (Krno 2007), (Krno & Holubec 2009), Krno & Žiak (2012).
Krno (1982) rozdelil povodie Ľupčianky na základné biotopy: krenál a ritrál. Pre pramennú oblasť uvádza charakteristické druhy: Capnia vidua, Arcynopteryx dichroa a Diura bicaudata. Ritrál delí ïalej na dve zóny a štyri podzóny. Zóna 1500 – 800 m n.m. s dominantnými druhmi Protonemura nimborum, Leuctra autumnalis, L.rauscheri a Isoperla sudetica. Druhy Leuctra inermis a Protonemura praecox uvádza ako dominantné pre druhú zónu
800 – 530 m n. m.
Krno (1984) na základe identity dominancie taxocenóz pošvatiek rozdelil povodie Belej na tri základné biotopy: kryál, ritrál a krenál. V kryáli (nad 1800 m n. m.) sa pošvatky nevyskytujú. Ritrál delí na tri zóny a štyri podzóny. Zónu vysokohorských potokov (2000 – 1500 m n. m.) charakterizuje početný výskyt druhov Protonemura brevistyla, Leuctra pusilla a L.rosinae. Charakteristickými dominantnými druhmi horských potokov (1500 – 800 m n. m.) sú Leuctra rauscheri, L.autumnalis, Isoperla sudetica a Protonemura montana. V zóne podhorských potokov (850 – 680 m n. m.) dominujú Protonemura praecox, Amphinemura sulcicollis a Leuctra inermis. Krenál delí na horské pramene s dominujúcimi druhmi Arcynopteryx dichroa a Amphinemura standfussi a podhorské pramene, pre ktoré uvádza dominantné druhy Protomenura auberti a Leuctra nigra.
Krno (1987) vypracoval dendrogram hierarchickej klasifikácie tečúcich vôd povodia horného Váhu na základe 10-ročných zberov makrozoobentosu. Autor diferencuje kryál (nad 1800 m n. m. v súčasnosti je presnejšie v súvislosti s povodím horného Váhu skôr používať termín nival (Krno et al. 2012) a ritrál (do 1800 m n.m.). Ritrál rozdeluje na tri podzóny epiritrálu (1800 – 750 m n. m.) a tri podzóny metaritrálu (pod 900 m n. m.). Spomedzi pošvatiek v epiritráli dominovali Protonemura auberti, Nemoura monticola, Leuctra atumnalis, L. rauscheri a Isoperla sudetica. Pre metaritrál uvádza charakteristické druhy Amphinemura sulcicollis, A. standfussi, Perlodes microcephala, Protonemura intricata a Leuctra digitata.
Predchadzajúce klasifikácie (Krno 1982, 1984, 1987) korešpondujú s tvrdením Illiesa (1952), ktorý tvrdí, že epiritrál obýva reofilná psychrostenotermná a polyoxybiotická glaciálna fauna, zatiaľ čo eurytermná postglaciálni imigranti a zmiešaná glaciálna fauna prevláda v metaritráli.
Krno (2003) uvádza úzku súvislosť medzi rozšírením taxocenóz pošvatiek a nadmorskou výškou, spádom a šírkou toku. Vymedzenie hraníc spoločenstiev nadmorskou výškou môžeme brať len orientačne a v úzkej korelácii s ostatnými abiotickými a biotickými faktormi. Toto potvrdzuje blízka príbuznosť spoločenstiev determinovaných z dvoch stanovíšť kaňonovitej doliny, ovplyvnených studenými krasovými podzemnými vodami, pod vrchom Siná v Rádovom potoku, v nadmorskej výške relatívne v nižších plochách 850 a 920 m a spoločenstiev vysokopoložených horských tokov povodia Demänovky, v nadmorskej výške nad 1300 m. Levadinov (1981), Ward (1985), Krno (2003) uvádzajú teplotu ako rozhodujúci faktor určujúci zonáciu tokov.
Vplyv karbonátového podložia však spôsobuje 7-násobne vyšší obsah Ca2+ v Rádovom potoku proti povrchovým vodám kryštalinika sledovaného povodia (Tereková 1993). Vysoký obsah vápnika pravdepodobne spôsobuje, napriek zatienenosti kaňonu, zvýšenú pokryvnosť machov (Palustriella commutata), ktorá je porovnateľná s pokryvnosťou machov na otvorených vysokopoložených horských tokoch (Slapania undulata, Hygrohypnum duriusculum). Výsledky analýz nám potvrdili vplyv faktoru pokryvnosť machov na spoločenstvá pošvatiek.
Pohľady na to, ktoré faktory hrajú rozhodujúcu úlohu pri zonácii tokov sa rôznia. Statzner (1987) poukazuje na 5 ekologických faktorov ovplyvňujúcich lotické ekosystémy: a/ teplotný režim, b/ prietokový režim a ním ovplyvnený charakter substrátu, jeho stabilita, procesy interakcií medzi vodným prostredím na jednej strane a organizmami, ovzduším, hyporeálom na strane druhej a turbidita vodného stĺpca, c/ svetelný režim, d/ živiny ovplyvňujúce primárnu produkciu, e/ prísun alochtónneho materiálu. Ako uvádzajú Krno et al. (1996), veľký význam pre existenciu bentickej fauny má kvalita substrátu, ktorej výskyt v rámci toku je ovplyvnený aj rýchlosťou prúdu. V povodí Demänovky sa na základe CCA analýz ukázali ako štatisticky najvýznamnejšie ekologické faktory, vplývajúce na rozšírenie pošvatiek, faktory typ substrátu, nadmorská výška a erózny index. Cummins (1969), Pennak (1971) uvádzajú substrát ako najdôležitejší ekologický faktor vplývajúci na rozšírenie spoločenstiev makrozoobentosu. Rovnako Allan (1975) tvrdí, že substrát je základný aspekt potočného habitatu odrážajúci rýchlosť prúdu, determinujúci úkryty, distribúciu potravy a iné.
Hrubozrný substrát balvany a skaly vytvára podľa Hynesa (1970) stabilnejšie podmienky pre rozvoj rozmanitejšej a početnejšej fauny ako jemnozrný substrát piesok a bahno. Druhovo najchudobnejším spoločenstvom pošvatiek povodia Demänovky bolo spoločenstvo Rakového podhorského toku na stanovišti v PR Jelšie v nadmorskej výške 700 m s piesočnato-bahnitým dnom. Toto zistenie korešponduje s pozorovaním, že stratocenózy na stabilnejšom substráte sú druhovo bohatšie (Deván 1994). Gurz & Wallace (1984) zistili, že hrubšie substráty sú fyzikálne stabilnejšie, ale menej vhodné na naplavenie jemnozrného detritu. Podľa nich je biologická stabilita spojená s fyzikálnou, preto druhy živiace sa nárastami alebo hrubým detritom sú frekventovanejšie na hrubších substrátoch. Rabeni & Minshall (1977), rovnako ako Williams (1977) poukázali na najbohatšie osídlenie hrubých štrkov (2,5 – 3,5 cm), pretože práve takýto substrát obsahuje veľké množstvo intersticiálnych priestorov, ktoré sú bohato zásobované detritom. Rovnako ako piesok a bahno, aj skaly a balvany sú osídlené chudobnejšie. S týmto tvrdením sa zhodujú aj údaje Helešica (1995), o najvyššej frekvencii druhov na skalnato-štrkovom substráte s vyššími hodnotami turbulencie prúdu a neutrálnou reakciou vody. Benke et al. (1988) definuje veľkosť, stabilitu a heterogenitu substrátu ako rozhodujúce vlastnosti pre štruktúru spoločenstva vodného hmyzu. Aj keď mnoho autorov uvádza substrát, ako primárny faktor určujúci štruktúru spoločenstva v rámci každej lokality, môže byť reakcia hmyzu na ten istý substrát modifikovaná ostatnými, najmä limitujúcimi environmentálnymi faktormi.
Krno (1982) udáva v povodí Ľupčianky najvyšší počet druhov pošvatiek (38) v oblastiach, ktoré ležia od 700 – 800 m n. m. Vo vyšších a v najmä nižších polohách počet druhov klesá. Podobnú situáciu môžem potvrdiť z povodia Demänovky, kde bol najvyšší počet druhov (38) rovnako v oblastiach od 700 – 900 m n. m. Obidve zistenia korešpondujú z podobnými súvislosťami zistenými všeobecne vo viacerých karpatských tokoch (Krno 2003).
Existujú dva základné prístupy ku klasifikácii tokov – zonačný, ktorý uvádzajú vyššie uvedení autori a kontinuálny (Vannote et al. 1980). Zo 46 nezávislých environmentálnych faktorov vplývajúcich na ekosystémy povodí až 63% tesne súvisí s hodnotou rádu toku. S rastom vodnatosti klesá prísun organického materiálu alochtónneho pôvodu, naopak stúpa podiel primárnej produkcie. Väčšina organickej hmoty je uložená v tokoch 1. až 3. rádu, metabolizovaná je však predovšetkým vo veľkých tokoch 7. až 9. rádu. S rastom prietoku stúpa podiel makrofýt, machov, rias a TOM. Klesá podiel CPOM, FPOM a UFPOM (Nainman et al. 1987). Podobné vzťahy ako medzi detritom, nárastmi a vodnatosťou tokov zaznamenali Krno et al. (1996) aj pre trofické skupiny makrozoobentosu. Abundancia trofických skupín makrozoobentosu s výnimkou drvičov vzrastá s vodnatosťou. Bruns et al. (1987) poukázali, že počet druhov zoškrabávačov a filtrátorov rastie s rádom toku. To však nezistili u zberačov, ktoré dominovali v stredných tokoch 4. až 6. rádu.
Z bentických spoločenstiev horských potokov uvádza Krno (1983) prevahu drvičov a relatívne vysoký podiel predátorov v riečke Ľupčianke, pričom pri prechode do podhorských tokov vzrastá podiel zoškrabávačov. Hustejší zápoj pobrežnej vegetácie na dolnom toku riečky spôsobuje opätovné zvýšenie podielu drvičov. Pre postupné zvyšovanie množstva jemného organického detritu od prameňa smerom k ústiu narastá aj podiel zberačov jemného organického materiálu. Ekologické postavenie vyjadrené na základe trofických skupín makrozoobentosu je determinované postavením tokov v riečnom kontinuu (Krno et al. 1996).
Povodie Demänovky je tvorené tokmi 1. až 4. rádu a v úplnom súlade s teóriou riečneho kontinua tu v spoločenstve pošvatiek výrazne prevládajú drviče (Nemouridae) a relatívne vysoké je zastúpenie predátorov (Perlodidae). V tokoch 4. rádu pozvolne narastá podiel zoškrabávačov (Brachyptera). Rozdiel zonálneho a kontinuitného pohľadu na hodnotenie vodných biotopov sa postupne prekonáva, Perry & Schaffer (1987) tvrdia, že gradient zmien taxonomického zloženia makrozoobentosu v tokoch je postupný, avšak spojený s náhlymi zmenami, ktoré sú sprevádzané zjavným zvýšením diverzity. Hildew et al. (1987) hovoria o diskontinuálnych zmenách ako o dôsledkoch náhlych zmien prúdu, prietoku a spádu toku (hydraulický stres).
Na erózne procesy odlesnených zjazdových tratí lyžiarskeho strediska Jasná pod Chopkom poukazuje Midriak (1993). Viacročnými meraniami zistil, že nechránený erodovaný povrch zjazdovej trate FIS sa zmyvom pôdy znižuje ročne o 4,6 – 7 mm oproti smrekovému lesu v rovnakých podmienkach 0,03 mm/rok, pričom intenzita pôdotvorných procesov je len pár stotín mm ročne. Unášané pôdne častice menia aj ekologické podmienky vodného prostredia toku. Nové ekologické podmienky sa odrážajú aj na druhovom zložení a zložení potravných gíld spoločenstva makrozoobentosu. Milner et al. (1981) a Rivier & Sequier (1985) tvrdia, že zvýšená sedimentácia spôsobuje kolmatáciu intersticiálnych priestorov a zanášanie povrchových biofilmov, čo výrazne negatívne ovplyvňuje štruktúru bentosu. Krno et al. (1995) zistili, že zvýšená erózia v povodí spôsobila obrusovanie biofilmov, zvýšený objem transportovaného organického materiálu (TOM), usadzovanie jemných sedimentov a kolmatáciu dna, výrazné zníženie indexu diverzity, abundancie makrozoobentosu a α-diverzity pošvatiek. V trofickej štruktúre došlo k výraznému poklesu zberačov zahrabaných v sedimentoch a zoškrabávačov. Celkový pokles abundancie makrozoobentosu je sprevádzaný nižšou abundanciou predátorov a zvýšeným podielom abundancie filtrátorov.
Najnápadnejší rozdiel medzi spoločenstvami makrozoobentosu oboch stanovíšť (2 a 10 – ovplyvné eróziou) je zastúpenie potravinovej gildy zberačov, ktorých podiel výrazne klesol v erodovanom potoku Priečne. V porovnaní s kontrolným stanovišťom, bolo na erodovanom toku (lok. 10) 10 krát menej zberačov, 5-násobne menej filtrátorov, naopak 12-násobne viac drvičov, 2-násobne viac predátorov a zoškrabávačov, pričom celková abundancia je približne rovnaká (Gašperčíková 1998).
Pri meraní jemného a ultrajemného usadeného materiálu sme zistili, že jeho celkové množstvo na m2 substrátu je pri oboch lokalitách rovnaké. Zásadný rozdiel však bol v pomere anorganickej a organickej zložky bentického jemného substrátu. Pri vyrovnanom neerodovanom toku bol pomer anorganiky k organike 1–1,5 ku 1. Pričom na erodovanom toku Priečne, anorganická zložka 6 – 13-krát prevyšovala organickú (Gašperčíková 1998). Krno et al. (1995) uvádzajú, že biomasa jemného usadeného organického detritu (FBOM) dosahuje maximá v dolných tokoch povodia Turca. Domnievame sa, že toto je spôsobené väčším množstvom intrersticiálnych priestorov substrátu ako i menším spádom tokov vyššieho rádu. V priemere 5-násobne väčšie množstvo transportovaného anorganického materiálu (TAM) na erodovanom toku Priečne oproti kontrole, spôsobuje väčšie usádzanie jemného anorganického materiálu, ktorý je ťažší, usádza sa prvý a upcháva intersticiálne priestory substrátu. Na základe vyššie uvedených zistení sa domnievame, že výrazne znížený podiel zberačov zahrabaných v sedimentoch je spôsobený upchávaním intersticiálnych priestorov substrátu anorganickým materiálom. Upchávanie intersticiálnych priestorov anorganickým materialom neumožňuje dostatočný prísun organického materiálu. Za primárnu príčinu nízkeho podielu zberačov z výnimkou rodu Leuctra (pravidelne unikajúci do hyporeálu) považujem preto práve dostupnosť potravy. Podobne klesá podiel fitrátorov na skalnatom substráte, ako dôsledok erózie anorganického material (Gašperčíková 1998).
Bruns et al. (1987) zaznamenali, že druhové bohatstvo zoškrabávačov je priamo závislé na biomase perifytonu a konštatujú. že pre zoškrabávačov je dôležitejšia rozmanitosť potravných zdrojov ako vlastný prietok. Zvýšený podiel zoškrabávačov na erodovanom toku Priečne je spôsobený 12-násobne väčším množstvom nárastov (Gašperčíková 1998). Gollday et al. (1992) uvádzajú, že pre eróziou narušené pôdy je typický rýchly proces nitrifikácie, ktorý produkuje H+ ióny, ktoré zvyšujú pH a prispievajú k zvýšenému stupňu mineralizácie. Vplyvom erózie pôdna voda prináša do toku väčšie množstvo nutrientov, hlavne vápenatých a draselnatých iónov. S väčším množstvom nutrientov a väčším osvetlením toku súvisí väčšie množstvo perifytonu v danom toku. Minshall et al. (1992), Krno et al. (1995) uvádzajú negatívny vplyv erózie na perifyton obrusovaním. Na erodovanom toku Priečne sme tento jav nepotvrdili, zrejme vďaka nižšiemu hydraulickému stresu. Jemné zbrusovanie mohlo na perifyton naopak pôsobiť ozdravujúco, pretože je vyššia pravdepodobnosť, že z nárastu uvoľňovali práve odumreté časti. Avšak algofágy zo skupiny pošvatiek – rod Brachyptera boli na erodovanom toku prakticky eliminované.
Druhové bohatstvo filtrátorov nie je závislé od absolútneho množstva potravy, ale od jeho plynulého prísunu a kvality (Bruns et al. 1987). Všeobecným indikátorom kvality potravy je percento organického materiálu. Znížený podiel hlavne TOM môže byť zvlášť kritickým pre filtrujúce Trichoptera, ktoré sa živia práve touto frakciou (Minshall et al. 1992). V zhode s týmito tvrdeniami je zaujímavý fakt, že 5-násobne menší podiel TOM ku TAM v porovnaní s kontrolným stanovišťom zhodne korešponduje s 5-násobne menším podielom filtrátorov (Gašperčíková 1998) v zastúpení potravných gíld makrozoobentosu erodovaného toku voči kontrole. Webster & Gollday (1984) uvádzajú, že pre erodované toky je charakteristické niekoľkonásobne väčšie množstvo TAM oproti TOM. Krno et al. (1995) zistili 4-násobne vyššie množstvo TAM ako TOM z časti Turca, podliehajúcemu erózii.
Gollday et al. (1992) uvádzajú, že abundancia drvičov v toku zostáva po odstránení lesa rovnaká, prejavujú sa však zjavné zmeny v štruktúre spoločenstva drvičov. Abundancia drvičov je najvyššia v tokoch 1. rádu (Krno 2003). Vysoký stav abundancie drvičov z pošvatiek reprezentovaný najmä rodmi Nemoura a Protonemura na erodovanom toku považujem v súlade s týmito tvrdeniami za pôvodný, eróziou nezmenený. Naopak, veľmi nízka abundancia drvičov na kontrolnom stanovišti neerodovaného toku Demänovka, bola zrejme spôsobená nižším pH vody. Bitušík & Koppová (1997) poukazujú na nárast stupeň acidifikácie v severných povodiach masívu Ďumbier, ktorý nastal koncom 90. rokov. Pri meraní pH kumulatívnych aj okamžitých zrážok v Demänovskej doline Tereková (1993) zistila ich výraznú aciditu (pH 4,7 – 5,0). Vplyvom erózie zvýšený prísun Ca2+, K+ a H+ do potoka Priečne eliminuje vplyv kyslých dažďov na pH erodovaného toku. Na kontrole úplne chýbal drvič Gammarus fossarum, ktorý bol naopak v erodovanom toku bol veľmi početný. Gammarus fossarum je výborným indikátorom acidifikácie, mizne z vôd s pH nižším ako 6,0 (Bitušík et al. 1989).
Významnými indikátormi kvality vodného prostredia sú Plecoptera. Na erodovanom toku Priečne bol zjavne vysoký počet zástupcov rodu Nemoura. Obdobný stav bol zistený na hornom toku Štrbského potoka, ktorý bol výstavbou novej diaľnice rovnako vystavený eróznemu tlaku (Krno 2000), ako aj na erodovanom toku Turčeka spolu rodom Leuctra (Krno et al. 1995). Ako veľmi dobrá metrika erózneho tlaku – LN_index (erózny index) na horské a podhorské potoky sa mi javí podiel zástupcov týchto dvoch rodov ku všetkým pošvatkám (Krno et al. 2015). Tento index dosahoval v povodí najvyššie hodnoty v dolných tokoch potoka Priečne (lyžiarsky terén) a Demänovky (polia).
Poďakovanie
Táto publikácia bola podporovaná grantom VEGA 1/0176/12, 1/0255/15.
Záver
Z 13 lokalít povodia Demänovky sme vo výskumnom období október 1995 – august 1997 determinovali 47 taxónov pošvatiek. Najpočetnejšie boli zastúpené rody Leuctra, Protonemoura a Isoperla. Počtom druhov, aj počtom jedincov prevládali na všetkých stanovištiach detritofágy, menej bolo predátorov a najmenej zoškrabávačov.
Pomocou permutačného testu CCA analýz sme z 15 environmentálnych faktorov vyselektovali 3 najvýznamnejšie, ktorých vplyv na celkovú ordináciu je štatisticky preukazný. Najdôležitejšie boli premenné percentuálne zastúpenie substrátu skaly, nadmorská výška a erózny index.
Na základe multivariačných analýz sme v povodí Demänovky vyčlenili 4 základné spoločenstvá patriace do dvoch základných skupín:
A – Spoločenstvá horských tokov 1, 2
B – Spoločenstvá podhorských tokov 3, 4
1. Spoločenstvá pošvatiek vysokopoložených horských tokov a tokov kaňonovitých dolín s úkazom teplotnej inverzie s dominantným druhom Leuctra pusilla a typickými Protonemura brevistyla, P. auberti.
2. Spoločenstvá pošvatiek vodnatejších horských tokov s charakeristickými druhmi Brachyptera starmachi, Protonemura nimborum, Leuctra rauscheri, Perlodes intricatus a Siphonoperla neglecta.
3. Spoločenstvá pošvatiek podhorských vodnatejších tokov s dominantným druhom Leuctra inermis a vlastnými druhmi spoločenstva Nemoura flexulosa, Perlodes microcephalus, Perla grandis a Siphonoperla torrentium.
4. Spoločenstvá pošvatiek podhorského pramenného toku s piesočnato-bahnitým dnom s dominantným druhom Nemurella pictetti a charakteristickým druhom Amphineura standfussi.
Vysoko položené bystriny majú nižšiu druhovú diverzitu, xenosaprobný stupeň, ktorý v podhorských tokoch prechádza do oligosaprobného.
Potok Priečne bol vystavený eróznemu tlaku, čo sa prejavilo výrazne vyšším transportom anorganického materiálu a následne vyšším podielom jemnejšieho substrátu (akál a piesok) usadeným na dne. V potravných funkčných skupinách makrozoobentosu v ňom bolo viac drvičov, predátorov a zoškrabávačov, a menej filtrátorov a zberačov.
Plecoptera sú významnými indikátormi kvality vodného prostredia. Ako dobrý indikátor zvýšenej erózie toku vychádza podiel počtu zástupcov rodu Nemoura a Leuctra ku všetkým pošvatkám. Navrhovaný index je vhodný pre horské a podhorské spoločenstvá karpatských tokov (ekoregión karpatikum).
Literatúra
Allan JD, 1975: The distributional ecology and diversity of benthic insects in Cement Creek, Colorado. Ecology, 56: 1040–1053.
Aubert J, 1959: Plecoptera. Fauna Helvetica, Lausanne, 1, 140 pp.
Balco M, 1977: Príspevok k hydrológii povrchových vôd Liptova. Liptov, 4: 5–57.
Benke AC, Hall CHAS, Hawkins CHP, Lowe-Mcconnell RH, Stanford JA, Suberkpp K & Ward JV, 1988: Bioenergetic consideretions in the analysis of stream ecosystems. J. N. Am. Benthol., 7: 480–502.
Beracko P, Lukáš J & Chvojka P, 2012: Časopriestorové zmeny spolčenstva potočníkov (Trichoptera) toku Demänovka, Folia faunistica Slovaca, 17 (1): 11–20.
Bitušík P, Krno I & Šporka F, 1989: Makrozoobentos dvoch ľadovcových jazier v Nízkych Tatrách. In: Bitušík, P. (ed) Stredné Slovensko 8, Prírodné Vedy, Osveta, Martin, p. 123–133.
Bitušík P & Koppová K, 1997: Macrozoobenthos of the glacial lakes in the Low Tatras (West Carpathians): Aquatic insects. Biológia (Bratislava), 52 (2): 227–232.
Braukmann U, 1987: Zoozonologische und saprobiologische Beitrage zur einer allgemeinen regionalen Bachtypologie. Archiv für Hydrobiol., Ergeb. der Limnol., 26: 355 pp.
Brink P, 1949: Studies on Swedish stoneflies. 0pusc. Ent. Suppl., 11: 1–250.
Bruns DA, Hale AB & Minshall GW, 1987: Ecological correlates of species richness in three guilds of lotic macroinvertebrates. Journal of Freashwater Ecology, 4: 163–175.
Berg K, 1948: Biological studies on the river Susaa. Fauna Limnol. Scandinavica, 4: 1–318.
Cernusca A, 1987: Wintersporterschliessungen und Naturschutz-Ergebnisse einer Studie im Auftrag des Europarates. Verh. Ges. Okol., 15: 173–181.
Cummins KW, 1969: The influence of substrate particle size on the microdistribution of stream macrobenthos. Hydrobiologia, 34: 145–181.
Čunderlíková B & Marhold K, 1984: Príspevok k poznaniu vegetácie lyžiarskych zjazdoviek vo Vysokých a Západných Tatrách. In: Zborník TANAP, 25, Martin, p. 89–116.
Deván P, 1994: Mayfly communities (Ephemeroptera) of the Myjava stream. Biologia (Bratislava), 49: 223–235.
Droppa A, 1957: Demänovské jaskyne. SAV Bratislava, 287 pp.
Droppa A, 1970: Výskum riečnych terás v zátopovej oblasti Liptovskej Mary. Liptov, 1: 7–34.
Droppa A, 1972: Geomorfologické pomery Demänovskej doliny. Slovenský kras (Liptovský Mikuláš), 10: 9–46.
Frutiger A & Imho A, 1997: Life cycle of Dinocras cephalotes end Perla grandis. In: Lanoldt P & Sartori M, (eds.), Ephemeroptera & Plecoptera, Mauron + Tinguely & Lachat sa, Fribourg, Switzerland, p. 34–43.
Frantz H, 1979: Okologie der Hochgebirge. Verl. Eugen Ulmer, Stuttgart, 495 pp.
Gašperčíková A, 1998: Analýza vplyvu vybraných ekologických faktorov na zloženie makrozoobentosu. Pošvatky (Plecoptera) povodia Demänovky. Prírodovedecká fakulta UK, 82 pp. Diplomová praca.
Gollday SW, Webster JR, Beneflied EF & Swank WT, 1992: Changes in stream stability following forest clearing as indicated by storm nutrient budgets. Arch. Hydrobiol., 90: 1–33.
Graf W, Lorenz AW, Tierno de Figueroa JM, Lucke MJ, López-Rodriguez MJ & Davies C, 2009: Distribution and ecological preferences of European freshwater organisms. Plecoptera. Pensoft, Moscow, 262 pp.
Gurtz ME & Wallace JB, 1984: Substrate medial response of stream invertebrates to disturbance. Ecology, 65: 1556–1559.
Hawkes HA, 1975: River zonation and classification. In: Whiton BA (ed.), River ecology, Blackwell Sientific Publ., Oxford, UK, p. 312–374.
Helan J, Kubíček, F, Losos B & Zelinka M, 1973: Production conditions in the trout brooks of the Beskydy mountains. Folia Fac. Sci. natur. Univ. Purkyn. Brun., Biol., 14: 1–105.
Helešic J, 1995: Pošvatky (Plecoptera) jako modelová skupina biologického modelování v tocích., Pŕírodovědecké fakulty M.U. v Brne, 120 pp. Dizertační práce.
Hildrew AG & Townsend CR, 1987: Organization in freshwater benthic communities. In: Gee JHR & Giller PS (eds.), Organozation of communities. Blackwell Scient. Publ., Oxford, p. 347–371.
Hrnčiarová T, 1996: Krajinnoekologické predpoklady pre návrh lyžiarskych zjazdových tratí. Ochrana prírody, Banská Bystrica, 14: 219–234.
Hynes HBN, 1961: The invertebrate fauna of a Welsh mountain stream. Arch. Hydrobiol., 57: 344–388.
Hynes HBN, 1970: The ecology of running waters. Liverpool, Univ. Press, 555 pp.
Illies J, 1952: Die Mölle. Faunistisch-ökologische Untersuchungen an einem Forellenbach im Lipper Bergland. Arch. Hydrobiol., 46: 424–612.
Illies J, 1961: Versuch einer allgemein biozönotischen Gleiderung der Fliesgewasser. Int. Rev. ges. Hydrobiol. Hydrog. 46: 205–213.
Illies J & Botosaneanu I, 1963: Problémes et metódes de la classification et de la zonation ecologique des eaux courantes, considerees surtout du point de vur faunistique. Int. Ver. Theor. Angew Limnol. Mitt., 12: 1–57.
Kocian Ľ, 1992: Vplyv lyžiarskej zjazdovky v Roháčoch na výskyt suchozemských stavovcov. Zborník prác o Tatranskom Národnom parku, 32: 363–376.
Krno I, 1978: Zoobentos rieky Revúcej a jej prítokov. Biologické práce SAV, Bratislava, 24, 63 pp.
Krno I, 1982: Štruktúra a dynamika makrozoobentosu rieky Ľupčianky a jej prítokov (Nízke Tatry). Biologické práce SAV, Bratislava, 28, 132 pp.
Krno I, 1983: Trofické skupiny makrozoobentosu v povodí rieky Ľupčianky. Biológia, 38: 145–148.
Krno I, 1984: Plecoptera des Einzugsgebietes des Flusses Belá. In: Ertl M (ed.), Limnologie des Flusses Belá. Práce Laboratória rybárstva a hydrobiológie, 4, p. 159–191.
Krno I, 1987: Classification of streams of the upper Váh River basin (West Carpathians). Acta Fac. Rer. natur. Univ. Comen., Zool., 29: 33–51.
Krno I, 2000: Makrozoobentos v povodí Bieleho Váhu, jeho pôvodnosť a prognóza jeho zmien. Acta Environmentalica Universitatis Comenianae, Bratislava, UK, 10: 197–205.
Krno I, 2003 : Distribution patterns and habitats of stoneflies in Slovakia. Research Update on Ephemeroptera and Plecoptera. – Perugia : University of Perugia, 349–356.
Krno I, 2007: Impact of human activities on stonefly (Insecta, Plecoptera) ecologiocal metrics in the Hron River (Slovakia). Biologia, 62 (4): 446–457.
Krno I, 2013: Pošvatky (Plecoptera): Determinačný kľúč pre hydrobiológov. Časť II. 1. vyd. – Bratislava: Výskumný ústav vodného hospodárstva, 64 pp.
Krno I & Holubec M, 2009: Effects of land use on stonefly bioassessment metrics. Aquatic Insects, 31 (Suppl. 1): 377–389.
Krno I, Šporka F, Tirjaková E & Bulánková E, 1995: Influence of the construction of the Turček reservoir on the organisms of the river bottom. Folia Fac. Sci. Nat. Univ. Masarykianae Brunensis, 91: 53–62.
Krno I, Šporka F, Tirjaková E, Bulánková E, Deván P, Degma P, Bitušík P, Kodada J, Pomichal R & Hullová E, 1996: Limnology of the Turiec river basin (West Carpathians, Slovakia). Biologia (Bratislava), 51 (Suppl.): 122 pp.
Krno I, Šporka F, Lánczos T & Štefková E, 2012: Vplyv prirodzených disturbancií na ekologický status tatranských bystrín na modelovom objekte pošvatiek (Plecoptera), 16. konferencia Slovenskej limnologickej spoločnosti a České limnologické společnosti. Bratislava: Slovenská limnologická spoločnosť. Jasná–Nízke Tatry, 25.–29.6.2012, p. 84–85,
Krno I, Šporka F, Lánczos T & Štefková E, 2015 Evaluation of deforestation influence caused by windstorm on the ecological status of the Carpathian streams by using Plecoptera assemblages as indicators, Appl. Limnol., (in press).
Krno I & Žiak M, 2012: Macrodistributions and microdistributions of stoneflies of calcareous submontane rivers of the West Carpathians, with different land cover. Aquatic Insects. 34 (1): 65–84.
Lacika J, 1992: Relief štátnej prírodnej rezervácie Demänovská dolina a niektoré problémy jej ochrany. Slovenský kras, 30: 89–102.
Levadinov VJ, 1981: Ekosystemy lososovych rek Dalnego vostoka. Trudy BPI. DVNC AN SSSR, Vladivostok, p. 3–21.
Lujkniš M (ed.), 1972: Slovensko 2. Príroda, Bratislava, Obzor, Bratislava, 917 pp.
Maitland PS, 1966: The fauna of the river Endrick. Studies on Loch Lomond., London, 2: 1–194.
Jakál J & Mazúr E, 1980: Atlas Slovenskej socialistickej republiky. SAV a Sloven. úrad a kartog., Bratislava, 296 pp.
Midriak R, 1993: Ochrana pôdy a krajinno-ekologická únosnosť územia Národného parku Nízke Tatry. Ochrana prírody, Banská Bystrica, 12: 9–53.
Milovanovič M, Ristič R, Radivojevič S, Nikcevič R & Malusevič I, 2007: Erosion control in ski areas. International Conference Erosion and Torrent Control as a Factor in Sustainable River Basin Management, 25–28 September, Belgrade.
Milner NJ, Sculling PA & Crisp DT, 1981: The effects of discharge on sediment dynamics and cosequent effects on invertebrates and salmonids in upland rivers. Appl. Ecol. 6: 153–200.
Mishall GW, Petersen RC, Bott TL, Cushing CE, Cummins KW, Vannote RL & Sedell JR, 1992: Stream ecosystem dynamics of the Salmon river, Idaho: an 8th-order system. J. N. Am. Benthol., 11: 111–137.
Naiman RJ, Melille JM, Lock MA & Ford TE, 1987: Longitudinal patterns of ecosystem processes and community structure in a subartic River continuum. Ecology, 68: 1139–1156.
Pennak RW, 1971: Toward a classification of lotic habitats. Hydrobiologia, 38: 321–334.
Perry JA & Schaeffer DJ, 1987: The longitudinal distribution of riverine benthos: A river discontinuum. Hydrobiologia, 148: 257–268.
Pielou EC, 1966: The measurement of diversity in different types of biological collections. J. Theor. Biol., 13: 131–144.
Rabeni CF & Minshall GW, 1977: Factors affecting microdistribution of stream bentic insects. Oikos, 26: 33–43.
Raušer J, 1964: Verbreitungsgeschichte der tschechoslowakischen Plecopteren – assoziationen. Gewäss. Abwäss. Dusseldorf, 34–35: 115–129.
Rivier B & Sequier J, 1985: Physical and biological effects of gravel extraction in river beds. In: Alabaster S (ed.), Habitat modification in freshwater fisheries. Butterworths, London, p. 131–146.
Shannon CE & Weaver W, 1949: The mathematical theory of communication. Univ. Illinois Press, Urbana.
Soldán T, Zahrádková S, Helešic J, Dušek L & Landa V, 1998: Distributional and quantitative patterns of Ephemeroptera and Plecoptera in the Czech Republic: A possibility of detection of long-term environmental changes of aquatic biotopes. Folia Fac. Sci. Nat Univ. Masarykianae Brunensis, Biologia, 98: 1–305.
Statzner B, 1987: Characteristic of Lotic Ecosystems and Consequences for Future Research Directions. Ecological Studies, 61: 365–390.
Šimo E, 1972: Povrchové vody. In: Lukniš M (ed): Slovensko 2, Príroda, Bratislava, Obzor, p. 283–342.
Štefanovič M, Milojevič M & Milovanovič M, 2007: Erosion process in ski centres of Serbia. Institute for development of water resources “Jaroslav Černi”, Beograd, www.http/ tucson.ars.ag.gov/isco/isco 15:1–6.
ter Braak CJF, 1986: Canonical correspodence analysis: a new eigtenvector technique for multivariate direct gradient analysis. Ecology, 67: 1167–1179.
Tereková V, 1993: Geochémia vôd Demänovského jaskynného systému a jeho ochrana. Ochrana prírody, Banská Bystrica, 12: 105–137.
Vannote RL , Minshall GW, Cummins KW, Sedell JR & Cushing CE, 1980: The river continuum concept. Can. J. Fish. Aquat. Sci., 37: 130–137.
Ward JV, 1985: Thermal characteristics of running waters. Hydrobiologia, 125: 31–46.
Webster JR & Golladay SW, 1984: Seston transport in streams at Coweeta Hydrologie Laboratory, North Carolina. Verh. Int. Ver. Limnol., 22: 1911–1919.
Williams DD, 1977: Substrate size selection by stream invertebrates and the influence of sand. Limnol. Oceanogr., 23: 1030–1033.
Zaťko M, 1980: Príspevok k teplote vody povrchových tokov niektorých pohorí Slovenska. Acta Fac. Rer. Nat. Univ Comenianae, Geographica, 18: 3–11.
Zwick P. 1996a: Capacity of discontinuous egg development and its importance for the geographic distribution of the warm water stenotherm, Dinocras cephalotes (Insecta: Plecoptera: Perlidae). Annls Limnol., 32: 147–160.
Zwick P, 1996b: Variable egg development of Dinocras spp. (Plecoptera: Perlidae) and the stonefly seed bank theory. Freshwater Biology, 35: 81–100.
Žiak M, 2013: Rozšírenie a diverzita pošvatiek (Plecoptera) Slovenska na základe vybraných enviromentálnych faktorov. Dizertačná práca, Prírodovedecká fakulta Univerzity Komernského, Bratislava, 334 pp.