Folia faunistica Slovaca 21 (1) 2016: –83
Príspevok k výskytu denných motýľov (Lepidoptera: Papilionoidea) okolia obcí východného Slovenska, časť 4 – Miroľa (Laborecká vrchovina)
Alexander Čanády
Ústav biologických a ekologických vied, Katedra zoológie,
Prírodovedecká fakulta UPJŠ, Šrobárova 2, SK – 041 54 Košice, Slovakia
[alexander.canady@upjs.sk, alexander.canady@gmail.com]
Abstract: The author investigated butterflies of superfamily Papilionoidea in the surrounding of village Miroľa (49°15’ N, 21°37’ E, altitude 390–530 m, Laborecká vrchovina Mts) during years 2014–2015. Totally were identified 62 butterfly species and 1 648 individuals belonging to 5 families. Among species were also recorded several species with different status of threatened of European and National importance (Iphiclides podalirius, Lycaena dispar, Phengaris arion, Brenthis ino, Melitaea phoebe, M. diamina and M. britomartis). According to the habitat preferences of butterflies were recorded: 15 ubiqvistic species, 29 mesophilic species (8 spieces: mesophil-1, 14 species: mesophil-2 and 7 species: mesophil-3), 13 xero-thermophilic species (5 species: xerotermophil-1 and 8 species: xerotermophil-2), 4 hygrophilous and 1 tyrphophilous species. Similarly, studied site represents a set of several microhabitats, which create favourable conditions for the survival of several species. The obtained data help to spread knowledge of butterflies in the territory of north-eastern Slovakia (Laborecká vrchovina Mts).
Key words: village Miroľa, north-eastern Slovakia, Laborecká vrchovina Mts., Lepidoptera.
Úvod
Poznatky o zložení lepidopterofauny z územia Laboreckej vrchoviny, podobne ako zo susediaceho orografického celku Ondavská vrchovina (porovnaj novšie údaje napr.: Čanády 2011, 2012, 2014, 2015) sú doposiaľ veľmi sporadické. Predovšetkým to boli výskumy v jej západnej časti (Hrubý 1964, Čaputa 1985, 1987, Jászay & Panigaj 1987, Panigaj & Reiprich 1998, Panigaj 1999a, b, Panigaj & Richter 2003, Macková 2014), východnej časti (Panigaj 1994, 2000, Šmajdová 2003, Bartušová & Panigaj 2004) či jej centrálnej časti (Kuruc 2003). Napriek tomu, že územie je málo preskúmané, predošlý výskum potvrdil predpoklad výskytu pozoruhodných druhov motýľov nielen pre oblasť východného Slovenska, ale aj s celoslovenským či európskym významom. Medzi takéto druhy patria predovšetkým Euphydryas maturna (Panigaj & Reiprich 1998, Panigaj 1999a) a Euphydryas aurinia (Macková 2014), Argynnis laodice (Panigaj 1999b), Carcharodus flocciferus, Parnassius mnemosyne a Cupido alcetas (Panigaj 1994, 2000), Hyponephele lycaon (Panigaj 2000, Kuruc 2003), Pseudopilotes vicrama a Pyrgus armoricanus (Panigaj 2000, Šmajdová 2003, Bartušová & Panigaj 2004), Neptis sappho (Macková in lit.), Melitaea didyma (vlastné nepubl. údaje., Macková in lit.).
Hlavným cieľom predkladaného príspevku je predovšetkým nadviazať na predchádzajúce výsledky získané na sledovanom území severovýchodného Slovenska, zmapovanie a doplnenie údajov o výskyte denných motýľov (Papilionoidea) so zameraním predovšetkým na intravilán a extravilán obce Miroľa. Údaje majú zároveň slúžiť ako podklad pre podrobnejšie vyhodnotenie lepidopterocenózy územia Laboreckej vrchoviny v budúcnosti.
Materiál a metódy
V období rokov 2014–2015 bol uskutočnený terénny výskum v okolí obce Miroľa s cieľom zistenia spoločenstiev motýľov s dennou aktivitou. Skúmané územie (DFS 6696, 49°19’48,7”N, 21°43’54,8”E, 390–530 m n. m.) je situované na severovýchode Slovenska (subprovincia Východné Karpaty) a orograficky patrí do celku Laborecká vrchovina (kód orografického celku – 750).
Faunistický prieskum bol realizovaný klasickými entomologickými metódami (pomocou entomologickej sieťky) alebo pozorovaním v priebehu mesiacov apríl až október na troch študijných plochách: B (B1, B2) a C. Na uvedených stanovištiach sa výskum motýľov uskutočňoval pozdĺž transektovej línie použitím transektovej metódy (línia B1, B2 a C), počas ktorej boli jedince odchytávané 2,5 metra na pravú a ľavú stranu pozdĺž transektu a pozorované jedince 5 metrov pred sebou (Pollard 1977). Zároveň boli jedince odchytávané a pozorované aj náhodne (A) pri prechode obcou alebo územím katastra obce (obr. 1). Počas tohto prechodu boli vyhľadávané okrem lúčnych porastov aj iné rozmanité typy krovín, zárastov na okraji lesa resp. priamo v lesnom poraste, okraje vodných či podmáčaných biotopov a vegetácie za účelom zaznamenania vzácnejších druhov, alebo druhov viazaných na špecifickejšie habitaty. Všetky letiace resp. sediace jedince boli determinované priamo v teréne, zároveň boli jedincom zotrené krídelné šupiny tak, aby nedošlo k poškodeniu krídel a aby sa predišlo ich opätovnému spočítaniu. Údaje o výskyte druhov boli zaznamenané do terénneho protokolu a len v nevyhnutnom prípade boli ťažko určiteľné druhy odoberané k ďalšiemu laboratórnemu spracovaniu a determinované pomocou určovacích kľúčov a atlasov (Jakšić 1998, Bělín 1999, Slamka 2004).
Odchytené druhy motýľov boli zaradené do príslušných čeľadí podľa najnovšej systematickej klasifikácie (Laštůvka & Liška 2011, Pastorális et al. 2013).
Na základe biotopovej väzby (Beneš et al. 2002), boli motýle rozdelené do piatich skupín: U: ubikvista: druh, schopný žiť na všetkých biotopoch, vrátane agrocenóz a ruderálov; M1: mozofil-1: druhy žijúce na otvorených biotopoch predovšetkým na mezofilných lúkach; M2: mezofil-2: druhy preferujúce rozhranie lesných a lúčnych biotopov, lesné lúky a svetliny a pod.; M3: mezofil-3: druhy žijúce v lesných biotopoch; X1: xerotermofil-1: druhy žijúce na otvorených xerotermných biotopoch, prevažne na nízko stebelných stepných trávnikoch a skalných stepiach; X2: xerotermofil-2: lesostepné a krovinové druhy; H: hygrofil: druhy žijúce na podmáčaných lúkach a slatinách (eutrofných mokradiach); T: tyrfofil: druhy oligotrofných mokradí, buď tyrfobiontné (žijúce len na rašeliniskách) alebo tyrfofilné druhy (preferujúce rašeliniská).
Podľa mobility boli motýle rozdelené do deviatich kategórii: 1: extrémne sedentárne; 2: veľmi sedentárne; 3: sedentárne 4: skôr sedentárne; 5: menej sedentárne; 6: ochotné rozptýlenia; 7: mobilné; 8: veľmi mobilné; 9: extrémne mobilné (Bartonova et al. 2014).
Druhy boli zároveň podľa klasifikácie Tischlera (1949) zaradené do piatich stupňov dominancie: eudominantné (Ed; D ˃ 10,1 %), dominantné (D; D = 5,1–10,0 %), subdominantné (SD; D = 2,1–5,0 %), recedentné (R; D = 1,1–2,0 %) a druhy subrecedentné (SR, D < 1,0 %).
Na porovnanie druhovej zhody (podobnosti) porovnávaných študijných plochách boli použité indexy identity: Jaccardov index (Ja) a Sörensenov index (Sö). Na vyjadrenie druhovej rozmanitosti boli vypočítané indexy diverzity: Shannon-Weaverov index diverzity a vyrovnanosti (H a J). Na vypočítanie príslušných indexov bol použitý štatistický program PAST verzia 2.71b (Hammer et al. 2001). Rovnako na porovnanie početnosti spoločných druhov v oboch študijných plochách bol použitý chí-kvadrát test (χ2) za použitia štatistického programu GraphPad Prism version 5.01 (GraphPad Software, Inc., San Diego, California, USA).
Biotopy a dátumy odchytov
Motýle boli zaznamenané počas 12 odchytových termínov na vybraných študijných plochách s cieľom pokryť čo najväčšie spektrum biotopov: 02.05.2014 – (A, B1, B2, C); 11.06.2014 – (A, B1, B2, C); 12.07.2014 – (D); 04.08.2014 – (A, B1, B2, C), 04.09.2014 – (A, B1, B2, C); 16.04.2015 – (A, B1, B2, C); 19.05.2015 – (A, B1, B2, C); 06.06.2015 – (A, B1, B2, C); 02.7.2015 – (A, B1, B2, C); 07.08.2015 – (A, B1, B2, C); 01.09.2015 – (A, B1, B2, C); 07.10.2015 – (A, B1, B2, C).
(A) – intravilán a extravilán obce: odchyty a pozorovania jedincov priamo v obci, alebo počas prechodu katastrom obce. Biotopy odchytov predstavovali zmes biotopov značne narušených ľudskou činnosťou predovšetkým: záhrady, ruderálne a krovinné spoločenstvá okrajov ciest, vegetácia pozdĺž „Mlynského“ potoka, mokriny, mezofilné lúky, okraje lesných porastov, agrocenózy s rôznou intenzitou využívania a podobne.
B(B1) – „PR Miroľská slatina“: Chránené územie je ukážkou vzácnych lúčnych a slatinných spoločenstiev. Na neveľkej ploche (0,97 ha) územia PR sa nachádza celá škála slatinných spoločenstiev, z ktorých dominujú ostrice (Carex ssp.), páperník širokolistý (Eriophorum latifolium), na najmokrejších častiach vachta trojlistá (Menyanthes trifoliata) a iné zákonom chránené rastliny. Vzhľadom na vzácny biotop a zákonom chránené územie, ako aj z ohľadom na ochranu fauny a flóry bol odchyt uskutočnený na okraji PR na ploche 0,12 ha (dĺžka x šírka línie: 230 x 5 m, Obr. 1). Stacionár bol kosený raz ročne mechanickým spôsobom v období letných mesiacov.
B(B2) – mezofilná lúka nachádzajúca sa nad PR Miroľská slatina: pokračovanie línie odchytu B1 (severozápadne od obce), uskutočnený transektovou metódou na ploche 0,12 ha (dĺžka x šírka línie: 230 x 5 m) okrajom mezofilnej lúky a dubovo-hrabového lesa (Obr. 1). Stacionár bol kosený raz ročne mechanickým spôsobom v období letných mesiacov.
(C) – lúka pod Jasenčíkom: mezofilná lúka severovýchodne od obce. Odchyt bol uskutočnený líniovou transektovou metódou na ploche 0,12 ha (dĺžka x šírka línie: 230 x 5 metrov, Obr. 1). Stacionár bol kosený raz ročne mechanickým spôsobom v období letných mesiacov.
Výsledky A Diskusia
Počas prieskumu lepidopterofauny v rokoch 2014–2015 bolo celkovo zaznamenaných 1 648 jedincov 62 druhov motýľov s dennou aktivitou (Papilionoidea) patriacich do 5 čeľadí.
Z celkového počtu pripadalo na študijnú plochu (B1) 507 jedincov patriacich k 48 druhom, na plochu (B2) 407 jedincov 43 druhov a na plochu (C) 505 jedincov 57 druhov motýľov. Zároveň bolo odchytených v katastri obce (študijná plocha A) ešte 229 jedincov patriacich 41 druhom (tabuľka 1).
Podľa biotopovej väzby motýľov bolo zaznamenaných 15 ubikvistických druhov, 29 mezofilných druhov (8 druhov: mezofil-1, 14 druhov: mezofil-2 a 7 druhov: mezofil-3), 13 xerotermofilných druhov (5 druhov: xerotermofil-1 a 8 druhov: xerotermofil-2), 4 hygrofilné druhy a 1 tyrfofilný až hygrofilný druh (obr. 2). Na základe vyššie uvedených údajov môžem konštatovať, že na skúmanej lokalite sa nachádza viacero mikrohabitatov, ktoré vytvárajú vhodné podmienky pre prežívanie viacerých druhov. Okrem ubikvistických druhov (24,2 %) schopných využívať a prežívať na viacerých typoch biotopov, prevažnú väčšinu predstavovali druhy mezofilné (46,2 %). Významnou zložkou, ktorá bola aj v zhode s vegetáciou typickou pre slatiny bolo aj zaznamenanie viacerých druhov, ktoré sú viazané na biotopy podmáčaných lúk až rašelinísk (tabuľka 1, obr. 2). Uvedené biotopy z hľadiska ich významu pre výskyt motýľov sú ešte viac zdôraznené z pohľadu ich ohrozenosti. Odchytené druhy boli zaradené aj do viacerých kategórii podľa Červeného zoznamu Slovenskej republiky (Kulfan & Kulfan 2001, tabuľka 1). Rovnako podľa Červeného zoznamu európskych motýľov (van Swaay et al. 2010) druh hnedáčik podunajský (Melitaea britomartis) je považovaný za takmer ohrozený a modráčik čiernoškvrný (Ph. arion) dokonca za ohrozený. Zároveň druhy L. dispar a Ph. arion patria medzi chránené druhy európskeho významu v zmysle vyhlášky Ministerstva životného prostredia Slovenskej republiky č. 24/2003 Z. z., ktorou sa vykonáva zákon č. 543/2002 Z. z. o ochrane prírody a krajiny v znení neskorších predpisov (Pastorális et al. 2013).
Na základe odchytených motýľov a ich početnosti boli porovnávané za účelom porovnania biotopov aj jednotlivé študijné plochy (B1, B2 a C), pre ktoré bolo zaznamenaných 34 spoločných druhov, z ktorých viaceré druhy (Maniola jurtina, Aphantopus hyperanthus, Coenonympha pamphilus, Thymelicus sylvestris, Leptidea juvernica, Boloria selene) mali zároveň eudominantné až subdominantné zastúpenie v spoločenstve denných motýľov na všetkých troch študijných plochách (Tabuľka 1). Viaceré druhy (napr. Erynnis tages, Thymelicus sylvestris, T. lineola, Hesperia comma, Lycaena alciphron, L. hippothoe, Phengaris arion, Polyommatus semiargus, P. icarus, Brenthis ino, Melitaea phoebe, M. diamina ako aj väčšina druhov podčeľade Satyrinae) patriace medzi sedentárne sú dôležitejším ukazovateľom kvality spoločenstva (Bartušová & Panigaj 2004, Bartonova et al. 2014). Ich význam je predovšetkým v ich viazanosti na habitaty určitej kvality, z ktorých sa veľmi nevzďaľujú a pri akejkoľvek zmene jeho kvality veľmi citlivo reagujú svojou početnosťou. Táto platnosť sa prejavila aj odchytom na študijnej ploche B2, ktorá bola priamym pokračovaním línie B1. Výsledky odchytu naznačili výraznejšie rozdiely v početnosti i druhovom zložení. Jedenásť druhov (C. palaemon, A. cardamines, P. brassicae, L. alciphron, B. ino, B. daphne, V. atalanta, N. c-album, A. iris a M. phoebe) boli zaznamenané len na ploche B1. Rovnako aj pre plochu B2 bol zaznamenaný špecifický výskyt šiestich druhov (I. podalirius, P. malvae, P. argus, A. aglaja, I. lathonia a B. dia). S výnimkou troch druhov (P. brassicae, I. lathonia a N. c-album), ktoré predstavujú mobilné druhy, patrili ostatné druhy medzi sedentárne alebo skôr sedentárne podľa stupnice mobility (Bartonova et al. 2014). Napriek tomu, o vysokej druhovej podobnosti oboch študijných plôch svedčia aj pomerne vysoké hodnoty Jaccardovho (Ja = 68,5 %) a Sörensenovho indexu (Sö = 81,3 %). Vypočítaním indexov diverzity bol zároveň potvrdený aj vysoký stupeň diverzity a vyrovnanosti (Tabuľka 2). Porovnaním spoločných druhov medzi oboma porovnávanými plochami boli potvrdené štatisticky vysoko významné rozdiely v ich početnosti (χ2 = 103,6; df = 36; p<0,0001).
Naopak, veľká podobnosť v mikrohabitate, spoločne zaznamenané druhy (tabuľka 1) spolu s hodnotami diverzity a vyrovnanosti (tabuľka 2), potvrdili, že študijná plocha C bola najviac podobná ploche B1 (Ja = 75,0 %, Sö = 85,7 %). Napriek tomu, porovnanie početnosti spoločných druhov ukázalo štatisticky významné rozdiely (χ2 = 76,95; df = 44; p<0,002). Z výsledkov vyplýva, že aj napriek blízkosti jednotlivých študijných plôch (B1, B2 a C) môžeme zaznamenať rozdiely v prítomnosti (neprítomnosti) ako aj početnosti predovšetkým u sedentárnych a habitatovo špecificky viazaných druhov.
Porovnaním odchytených druhov v jednotlivých čeľadiach boli zistené nasledujúce odlišnosti v ich početnosti. Čeľaď Papilionidae bola zastúpená opäť dvomi druhmi v jarných a letných mesiacoch, čo je v zhode aj s predošlými údajmi (Čanády 2011, 2012, 2014, 2015). Vo vyššie uvedených prácach (t.j. v okolí skúmaných obcí Ondavskej vrchoviny), ani v okolí obce Miroľa (Laborecká vrchovina) ich celková početnosť nebola vysoká, ale ukázala, že oblasť severovýchodného Slovenska je tvorená habitatmi vytvárajúcimi vhodné podmienky na prežívanie viacerých druhov motýľov. Rovnako, druhy boli zaznamenané aj mimo študijných plôch pri prechode katastrom obce a bola pozorovaná ich veľmi dobrá mobilita (Beneš et al. 2002, Slamka 2004), aj keď patria skôr medzi sedentárne druhy (Bartonova et al. 2014). Potvrdenie výskytu ďalšieho druhu Parnassius mnemosyne v okolí obce Duplín (Čanády 2012) ako aj v okolí kopca Gazdoráň a v údolí pozdĺž potoka Chotinky (Panigaj 2000), naznačuje, že sa nedá celkom vylúčiť prítomnosť druhu na vhodných stanovištiach či už v okolí obce Miroľa, alebo na väčšom území Ondavskej a Laboreckej vrchoviny.
Čeľaď Hesperiidae bola zastúpená siedmimi druhmi, pričom najpočetnejší bol výskyt Thymelicus sylvestris s celkovým počtom 73 zaznamenaných jedincov a subdominantným výskytom (4,4 %). Výskyt druhu na uvedených stanovištiach bol aj v zhode s jeho biotopovou väzbou, keďže druh patrí do skupiny mezofil-2, teda druh preferujúci rozhranie lesných a lúčnych biotopov (Beneš et al. 2002, Slamka 2004). Okrem dominantného zastúpenia T. sylvestris, patrili medzi ďalšie druhy so subdominantným zastúpením aj T. lineola (2,0 %). Ostatné druhy mali len recedentné až subrecedentné zastúpenie. Pri porovnaní výskytu na jednotlivých stanovištiach (B1 a C) bolo zaznamenaných 6 spoločných druhov (Tabuľka 1), so štatisticky nevýznamným rozdielom v ich početnosti (χ2 = 7,62; df = 5; p = 0,179).
Z čeľade Pieridae bolo zaznamenaných osem druhov, ktorých odchyt bol potvrdený počas celého vegetačného obdobia až do neskorej jesene. Veľký podiel v odchytoch mali mlynáriky rodu Leptidea a Pieris, tvoriace stálu zložku fauny motýľov okolia obce Miroľa. Výsledky sú v zhode aj s údajmi zo širšieho okolia sledovaného územia (cf. Čanády 2011, 2012, 2014, 2015) a zároveň aj s ich habitatovými nárokmi a mobilitou (Beneš et al. 2002, Slamka 2004, Bartonova et al. 2014). Pre obe skúmané plochy (B1 a C) bolo zaznamenaných 8 spoločných druhov (Tabuľka 1) bez štatisticky významného rozdielu v ich početnosti (χ2 = 6,78; df = 7; p=0,453). Medzi subdominantné druhy patrili Leptidea juvernica (3,5 %) a L. sinapis (2,5 %) a z druhov rodu Pieris len P. rapae mal početnejšie zastúpenie v lepidopterocenóze (2,9 %).
Pre čeľaď Lycaenidae bolo zaznamenaných 13 druhov, pričom 9 druhov (Tabuľka 1) bolo spoločných pre obe lokality (B1 a C) bez štatisticky významného rozdielu v početnosti (χ2 = 15,20; df = 8; p = 0.06). Dominantné zastúpenie mali predovšetkým ohniváčiky rodu Lycaena (L. hippothoe, L. tityrus a L. virgaureae). Naopak, prekvapujúci bol pomerne malý výskyt takých druhov ako sú C. argiades (1,9 %) a P. icarus (1,5 %), ktoré sú veľmi dobre adaptované na najrôznejšie typy otvorených bezlesných biotopov vrátane poľnohospodárskej krajiny, intravilánov miest a obcí, ruderálnych stanovíšť, suchších zošliapaných lúk a poľných ciest (Beneš et al. 2002, Slamka 2004). Z faunistického a ochranárskeho hľadiska boli významné potvrdenia výskytu druhov L. dispar a Ph. arion, ktoré patria medzi chránené druhy európskeho významu.
Druhovo najpočetnejšou skupinou motýľov bola čeľaď Nymphalidae s 32 zaznamenanými druhmi a ich rôznym výskytom na jednotlivých študijných plochách. Medzi najpočetnejšie druhy patrili na oboch plochách najmä zástupcovia podčeľade Satyrinae (M. jurtina, C. pamphilus, M. galathea, A. hyperanthus, M. dryas a C. glycerion). Spoločných pre obe lokality bolo 22 druhov (Tabuľka 1), s vysoko významným rozdielom v ich početnosti (χ2 = 45,02; df = 21; p = 0,002). Významné boli odchyty a potvrdenia výskytu viacerých druhov (Argynnis niobe, Brenthis ino, Melitaea phoebe, M. diamina a M. britomartis), ktoré sú uvedené aj v Červenom zozname Slovenskej republiky so súčasným statusom zraniteľnosti až ohrozenosti.
Rozdielne výsledky početnosti jednotlivých druhov na sledovanom území ako aj na jednotlivých stanovištiach sú dôkazom toho, že na zhodnotenie reálneho stavu je pri vyhodnocovaní nutné zohľadňovať aj zloženie druhového spektra, na čo dobre slúžia indexy diverzity (pozri porovnanie vyššie). Bartušová & Panigaj (2004) vo svojej práci o vplyve obhospodarovania lúčnych biotopov na spoločenstvá denných motýľov potvrdili, že popis spoločenstva založený len na počte vyskytujúcich sa druhov ešte nič nehovorí o jeho diverzite. Diverzita zahŕňa okrem druhového bohatstva aj dôležitý aspekt numerickej štruktúry, ktorý predstavuje bežnosť a vzácnosť druhov.
Na základe získaných údajov o spoločenstve denných motýľov okolia obce Miroľa, sa dá konštatovať, že v jej okolí sa v súčasnosti nachádza pomerne dobre zachované druhové spektrum a vyrovnané spoločenstvo motýľov. Zároveň, potvrdenie výskytu vzácnejších druhov s celoeurópskym významom (I. podalirius, P. machaon, L. dispar, L. alcipron, Ph. arion, Brenthis ino, Melitaea phoebe, M. diamina a M. britomartis) ako aj prítomnosť vzácnej flóry PR Miroľská slatina len zdôrazňujú ochranu daného územia spojenú s vhodným manažmentom obhospodarovania lúčnych a lesných ekosystémov. Pri kosbe na kvetnatých lúkach nikdy nekosiť celú lúku súčasne, ale postupovať v mozaikách, pruhoch, ponechávať nekosené okraje, ktoré môžu byť následne skosené pri ďalšej kosbe o niekoľko dní neskôr, alebo až v nasledujúcom roku. Takým spôsobom by sa aspoň čiastočne priblížilo k tradičnému hospodáreniu z minulosti, kedy nikdy nedochádzalo k pokoseniu celých plôch naraz. Naopak, v súčasnosti môže veľkoplošné kosenie spojené priamo s mulčovaním v dobe plného letu niektorých druhov pripraviť nielen imága o zdroje nektáru, ale aj priame zničenie vajíčok a ďalších vývojových štádií. Lúky tiež nehnojiť, neodvodňovať a snažiť sa o obnovu druhovo bohatých stanovíšť. Na druhej strane, je ale možné drobnými mechanickými narušeniami vegetačného krytu (prejazdom vozidlom, pastvou a podobne) obohatiť podmienky stanovištia, čím sa podporí výskyt vzácnejších pastvinových druhov. Dôležité je aj zamedzenie sukcesie mezofilných ruderálov vyrezávaním krovín, spásaním a dokonca aj riadeným vypaľovaním. Osobitnú pozornosť si vyžaduje najmä územie ohraničené PR Miroľská slatina s výskytom slatinno-rašelinných spoločenstiev, pre ktorú je najvhodnejšia predovšetkým ručná kosba raz v roku buď skoro na jar v máji, alebo až neskoro na jeseň pri zachovaní nepokosených okrajov. Zároveň dohliadať aby nedošlo k odvodňovaniu územia (Beneš et al. 2002, Bartušová & Panigaj 2004).
Poďakovanie
Moje poďakovanie patrí doc. Dr. Ľ. Panigajovi za pomoc pri determinácii ťažšie určiteľných druhov a cenné pripomienky k rukopisu. Rovnako sa chcem poďakovať všetkým obyvateľom obce za trpezlivosť a ústretovosť počas výskumu.
Literatúra
Bartonova A, Benes J. & Konvicka M. 2014: Generalist-specialist continuum and life history traits of Central European butterflies (Lepidoptera) – are we missing a part of the picture? European Journal of Entomology, 111 (4): 543–553.
Bartušová Z & Panigaj Ľ, 2004: Vplyv obhospodárovania lúčnych porastov na štruktúru cenóz denných motýľov (Lepidoptera: Zygaenoidea, Hesperioidea et Papilionoidea). Ochrana prírody, 23: 253–264.
Beneš J, Konvička M, Dvořák J, Fric Z, Havelda Z, Pavlíčko A, Vrabec V & Wiedenhoffer Z, (eds.) 2002: Motýli České republiky: Rozšírení a ochrana I., II. 857 pp.
Bělín V, 1999: Motýli České a Slovenské republiky aktivní ve dne. Kabourek. Zlín. 95 pp., 43 tab.
Čanády A, 2011: Príspevok k výskytu denných motýľov (Hesperioidea, Papilionoidea) okolia obcí východného Slovenska, časť I. – Duplín (Ondavská vrchovina). Folia faunistica Slovaca, 16 (2): 79–83.
Čanády A, 2012: Príspevok k faunistike denných motýľov (Lepidoptera: Rhopalocera) z východného Slovenska za roky 2008–2011. Folia faunistica Slovaca, 17 (2): 151–157.
Čanády A, 2014: Príspevok k výskytu denných motýľov (Lepidoptera: Papilionoidea) okolia obcí východného Slovenska, časť 2 – Potoky (Ondavská vrchovina). Folia faunistica Slovaca, 19 (3): 251–260.
Čanády A, 2015: Príspevok k výskytu denných motýľov (Lepidoptera: Papilionoidea) okolia obcí východného Slovenska, časť 3 – Tokajík (Ondavská vrchovina). Folia faunistica Slovaca, 20 (1): 95–104.
Čaputa A, 1985: Inventarizačný výskum ŠPR Mokré lúky pod Čertižným. Textová časť. Manuskript. Správa CHKO Východné Karpaty, Humenné, 12 pp.
Čaputa A, 1987: Inventarizačný výskum ŠPR Haburské rašelinisko. Textová časť. Manuskript. Správa CHKO Východné Karpaty, Humenné, 6 pp.
Hammer Ø, Harper DAT & Ryan PD, 2001: PAST: Paleontological statistics software package for education and data analysis, Palaeontologia Electronica, 4: 9pp.
Hrubý K, 1964: Prodromus Lepidopter Slovenska. Vydavateľstvo SAV, Bratislava, 962 pp.
Jakšić NP, 1998: Male genitalia of butterflies on Balkan Peninsula with a check-list (Lepidoptera: Hesperioidea and Papilionoidea). Bratislava, Slovakia, 144 pp.
Jászay T. & Panigaj Ľ, 1987: Niekoľko poznámok k prieskumu motýľov (Lepidoptera) severovýchodnej časti okresu Svidník a k problematike entomologických výskumov. Prehľad odborných výsledkov, X. Východoslovenský TOP (Krajná Bystrá 1986), p. 81–90.
Kulfan M. & Kulfan J, 2001: Červený (ekosozologický) zoznam motýľov (Lepidoptera) Slovenska. 134–137 pp. In: Baláž D., Marhold K. & Urban P. (eds.), 2001: Červený zoznam rastlín a živočíchov Slovenska. Ochrana Prírody, 20 (Suppl.), 160 pp.
Kuruc Š. 2003:Distribúcia heliofilných motýľov (Lepidoptera: Hesperioidea a Papilionoidea) v centrálnej časti Laboreckej vrchoviny. Rigorózna práca. Deponovaná na PF UPJŠ v Košiciach. Košice. 23 pp.
Laštůvka Z. & Liška J, 2011: Komentovaný seznam motýlů České republiky. Biocont Laboratory. Brno. 48 pp.
Macková A, 2014: Nález hnedáčika chrastavcového (Euphydryas aurinia) na Slovensku po 13 rokoch. Pp. 135–137. In: Manko P. & Baranová B. (eds.): Zborník príspevkov z vedeckého kongresu „Zoológia 2014“, 19. Feriancove dni. Vydavateľstvo PU, Prešov.
Panigaj Ľ, 1994: Výskyt niektorých teplomilných druhov motýľov (Lepidoptera) v Bukovských vrchoch. Zborník Východoslovenského múzea v Košiciach, Prírodné vedy, 35: 191–192.
Panigaj Ľ, 1999a: Pozoruhodný nález Euphydryas maturna L. (Lepidoptera: Nymphalidae) na východnom Slovensku. Natura Carpatica, 40: 231–234.
Panigaj Ľ, 1999b: Poznámka k rozšíreniu Argyronome laodice Pall. (Lepidoptera: Nymphalidae) na Slovensku. Natura Carpatica, 40: 235–238.
Panigaj Ľ, 2000: Motýle Národného parku Poloniny. Štátna ochrana prírody SR, Banská Bystrica a Správa Národného parku Poloniny, Snina, 111 pp.
Panigaj Ľ. & Reiprich A, 1998: Niekoľko pozoruhodných nálezov motýľov (Lepidoptera) na východnom Slovensku. Entomofauna Carpatica, 10 (3): 85–90.
Panigaj Ľ. & Richter I. 2003: Poznámky k faune motýľov (Lepidoptera) západnej časti CHKO Východné Karpaty. Prehľad odborných výsledkov, XXVII. Východoslovenský TOP (Vyšná Pisaná 2003). Prešov, p. 60–75.
Pastorális G, Kalivoda H & Panigaj Ľ, 2013: Zoznam motýľov (Lepidoptera) zistených na Slovensku. Folia faunistica Slovaca, 18 (2): 101–232.
Pollard E, 1977: A method for assesing changes in the abundance of butterflies. Biological Conservation, 12: 115–134.
Slamka F, 2004: Die Tagfalter Mitteleuropas – östliche Teil. Bestimmung Biotope und Bionomie Verbreitung Gefährdung. Bratislava, 288 pp.
Šmajdová Z, 2003: Vplyv obhospodarovania lúčnych porastov na štruktúru cenóz denných motýľov. Diplomová práca. Deponovaná na PF UPJŠ v Košiciach. Košice, 78 pp.
Tischler W, 1949: Grundzüge der terrestrischen Tierökologie. Braunschwieg, Friedr. Vieweg, 219 pp.
Van Swaay C, Cuttelod A, Collins S, Maes D, López Munguira M, Šašić M, Settele J, Verovnik R, Verstrael T, Warren M, Wiemers M. & Wynhof I 2010: European red list of butterfies. Luxembourg: Publications office of the European Union, 48 pp.
Wahlberg N, Leneveu J, Kodandaramaiah U, Peña C, Nylin S, Freitas A.V.L. & Brower A.V.Z, 2009: Nymphalid butterflies diversify following near demise at the Cretaceous/Tertiary boundary. Proceedings of the Royal Society B, 276: 4295–4302.
Appendix
Prehľad motýľov podľa termínov odchytu a lokalít:
• čeľaď Vidlochvostovité (Papilionidae) – 2 spp. (12 ex.):
Iphiclides podalirius – 7 ex.: 02.05.2014 – 3 ex. (A); 04.08.2014 – 1 ex. (C); 06.06.2015 – 1 ex. (C); 07.08.2015 – 2 ex. (B2).
Papilio machaon – 5 ex.: 02.05.2014 – 1 ex. (A), 1 ex. (B1); 19.05.2015 – 1 ex. (B2); 07.08.2015 – 1 ex. (A); 01.09.2015 – 1 ex. (B2).
• čeľaď: Súmračníkovité (Hesperidae) – 7 spp. (172 ex.):
Erynnis tages – 17 ex.: 02.05.2014 – 3 ex. (B1), 1 ex. (B2), 1 ex. (C); 06.06.2015 – 1 ex. (B1); 04.08.2014 – 1 ex. (C); 19.05.2015 – 2 ex. (B1), 2 ex. (B2), 3 ex. (C); 06.06.2015 – 1 ex. (C); 07.08.2015 – 2 ex. (C).
Pyrgus malvae – 8 ex.: 02.05.2014 – 3 ex. (C); 19.05.2015 – 1 ex. (B2), 1 ex. (C); 06.06.2015 – 3 ex. (C).
Carterocephalus palaemon – 13 ex.: 02.05.2014 – 4 ex. (A), 3 ex. (C); 19.05.2015 – 1 ex. (A), 1 ex. (B1), 4 ex. (C).
Thymelicus lineola – 33 ex.: 12.07.2014 – 15 ex. (B1), 10 ex. (C); 04.08.2014 – 2 ex. (C); 07.08.2015 – 1 ex. (A), 1 ex. (B1), 1 ex. (B2), 3 ex. (C).
Thymelicus sylvestris – 73 ex.: 12.07.2014 – 4 ex. (B1), 2 ex. (B2), 8 ex. (C); 02.07.2015 – 25 ex. (B1), 17 ex. (B2), 15 ex. (C); 07.08.2015 – 1 ex. (A), 1 ex. (B1).
Hesperia comma – 16 ex.: 04.08.2014 – 1 ex. (B1), 1 ex. (B2); 07.08.2015 – 5 ex. (A), 5 ex. (B1), 2 ex. (B2), 2 ex. (C).
Ochlodes sylvanus – 12 ex.: 11.06.2014 – 1 ex. (A), 1 ex. (B1), 1 ex. (B2), 1 ex. (C); 04.09.2014 – 2 ex. (B1); 06.06.2015 – 1 ex. (C); 02.07.2015 – 2 ex. (C); 07.08.2015 – 1 ex. (B2), 01.09.2015 – 1 ex. (B1), 1 ex. (B2).
• čeľaď Mlynárikovité (Pieridae) – 8 spp. (214 ex.):
Leptidea sinapis – 41 ex.: 02.05.2014 – 2 ex. (A), 2 ex. (B1), 1 ex. (C); 12.07.2014 – 2 ex. (B1), 2 ex. (B2); 04.08.2014 – 1 ex. (B1), 1 ex. (B2); 04.09.2014 – 1 ex. (A), 1 ex. (C); 16.04.2015 – 1 ex. (C); 19.05.2015 – 1 ex. (B1), 1 ex. (C); 06.06.2015 – 1 ex. (B1); 02.07.2015 – 3 ex. (B1), 5 ex. (B2), 4 ex. (C); 07.08.2015 – 1 ex. (A), 3 ex. (B2), 4 ex. (C); 01.09.2015 – 1 ex. (A), 2 ex. (B1), 1 ex. (B2).
Leptidea juvernica – 58 ex.: 02.05.2014 – 6 ex. (B1), 5 ex. (B2), 5 ex. (C); 12.07.2014 – 4 ex. (B1), 3 ex. (B2), 3 ex. (C); 04.09.2014 – 1 ex. (B1), 1 ex. (B2); 19.05.2015 – 6 ex. (B1), 7 ex. (B2), 4 ex. (C); 06.06.2015 – 3 ex. (B2), 2 ex. (C); 02.07.2015 – 3 ex. (B1), 3 ex. (C); 07.08.2015 – 2 ex. (B1).
Anthocharis cardamines – 9 ex.: 02.05.2014 – 2 ex. (A), 2 ex. (B1), 4 ex. (C); 19.05.2015 – 1 ex. (C).
Pieris brassicae – 4 ex.: 02.05.2014 – 1 ex. (B1); 02.07.2015 – 2 ex. (C); 01.09.2015 – 1 ex. (C).
Pieris rapae – 48 ex.: 02.05.2014 – 1 ex. (A), 1 ex. (B1); 04.08.2014 – 2 ex. (A), 1 ex. (C); 04.09.2014 – 2 ex. (A), 4 ex. (B1); 02.07.2015 – 2 ex. (B1), 5 ex. (B2), 5 ex. (C); 07.08.2015 – 6 ex. (A), 4 ex. (B1), 4 ex. (B2), 1 ex. (C); 01.09.2015 – 6 ex. (A), 3 ex. (B1); 07.10.2015 – 1 ex. (B1).
Pieris napi – 31 ex.: 02.05.2014 – 5 ex. (A), 1 ex. (B2), 2 ex. (C); 11.06.2014 – 2 ex. (A); 12.07.2014 – 1 ex. (B1), 1 ex. (C); 04.09.2014 – 1 ex. (B1); 19.05.2015 – 1 ex. (A), 1 ex. (C); 06.06.2015 – 1 ex. (C); 02.07.2015 – 1 ex. (B2), 1 ex. (C); 07.08.2015 – 2 ex. (A), 3 ex. (B1), 1 ex. (B2), 1 ex. (C); 01.09.2015 – 2 ex. (A), 2 ex. (B1); 07.10.2015 – 2 ex. (B2).
Colias hyale – 9 ex.: 04.09.2014 – 1 ex. (B2), 1 ex. (C); 06.06.2015 – 1 ex. (B2); 07.08.2015 – 2 ex. (A); 01.09.2015 – 2 ex. (B1), 1 ex. (B2); 1 ex. (B2).
Gonepteryx rhamni – 14 ex.: 02.05.2014 – 2 ex. (A), 1 ex. (B1), 1 ex. (B2); 04.08.2014 – 1 ex. (A), 2 ex. (B2), 1 ex. (C); 16.04.2015 – 1 ex (C); 19.05.2015 – 1 ex. (C); 07.08.2015 – 2 ex. (A), 2 ex. (B1).
• čeľaď Ohniváčikovité (Lycaenidae) – 13 spp. (228 ex.):
Lycaena phleas – 2 ex.: 06.06.2015 – 1 ex. (B1), 01.09.2015 – 1 ex. (B2).
Lycaena dispar – 16 ex.: 12.07.2014 – 1 ex. (C); 04.09.2014 – 1 ex. (B2); 06.06.2015 – 1 ex. (A); 02.07.2015 – 1 ex. (A), 3 ex. (B1), 1 ex. (B2), 1 ex. (C); 01.09.2015 – 4 ex. (B1), 1 ex. (B2), 2 ex. (C).
Lycaena virgaureae – 25 ex.: 11.06.2014 – 1 ex. (A); 12.07.2014 – 1 ex. (A), 1 ex. (B2); 04.08.2014 – 1 ex. (B1), 1 ex. (B2); 02.07.2015 – 1 ex. (B1), 6 ex. (B2), 2 ex. (C); 07.08.2015 – 1 ex. (A), 1 ex. (B2), 1 ex. (C); 01.09.2015 – 5 ex. (B1), 3 ex. (B2).
Lycaena tityrus – 35 ex.: 02.05.2014 – 2 ex. (B2); 04.08.2014 – 3 ex. (B1), 3 ex. (B2); 19.05.2015 – 4 ex. (B2), 1 ex. (C); 06.06.2015 – 3 ex. (C); 07.08.2015 – 3 ex. (A), 5 ex. (B1), 9 ex. (B2), 2 ex. (C).
Lycaena alciphron – 16 ex.: 11.06.2014 – 4 ex. (A), 1 ex. (B1), 9 ex. (C); 02.07.2015 – 2 ex. (B1).
Lycaena hippothoe – 51 ex.: 11.06.2014 – 7 ex. (A), 27 ex. (B1), 2 ex. (B2), 9 ex. (C); 02.07.2015 – 3 ex. (B1), 2 ex. (B2), 1 ex. (C).
Thecla betulae – 3 ex.: 04.09.2014 – 1 ex. (B2); 07.08.2015 – 1 ex. (C); 01.09.2015 – 1 ex. (B1).
Cupido argiades – 31 ex.: 02.05.2014 – 1 ex. (C); 12.07.2014 – 2 ex. (B1), 4 ex. (C); 04.08.2014 – 1 ex. (B2); 04.09.2014 – 3 ex. (B2); 19.05.2015 – 1 ex. (C); 02.07.2015 – 1 ex. (B1), 3 ex. (B2); 07.08.2015 – 1 ex. (A), 6 ex. (B2), 2 ex. (C); 01.09.2015 – 1 ex. (A), 1 ex. (B1), 3 ex. (B2), 1 ex. (C).
Celastrina argiolus – 1 ex.: 01.09.2015 – 1 ex. (A).
Phengaris arion – 6 ex.: 12.07.2014 – 4 ex. (C); 04.08.2014 – 1 ex. (A); 02.07.2015 – 1 ex. (C).
Plebejus argus – 14 ex.: 11.06.2014 – 5 ex. (C); 12.07.2014 – 1 ex. (C); 04.08.2014 – 1 ex. (C); 07.08.2015 – 3 ex. (B2), 3 ex. (C); 01.09.2015 – 1 ex. (B2).
Polyommatus semiargus – 3 ex.: 11.06.2014 – 1 ex. (B1), 1 ex. (C); 06.06.2015 – 1 ex. (B2).
Polyommatus icarus – 25 ex.: 11.06.2014 – 1 ex. (C); 06.06.2015 – 1 ex. (B1); 02.07.2015 – 1 ex. (A); 07.08.2015 – 1 ex. (A), 1 ex. (B1), 2 ex. (B2), 2 ex. (C); 01.09.2015 – 5 ex. (A), 7 ex. (B1), 3 ex. (B2), 1 ex. (C).
• čeľaď Babôčkovité (Nymphalidae) – 32 spp. (1022 ex.):
Argynnis paphia – 4 ex.: 04.08.2015 – 1 ex. (C); 07.08.2015 – 1 ex. (A), 2 ex. (C).
Argynnis aglaja – 2 ex.: 12.07.2014 – 1 ex. (A); 04.08.2014 – 1 ex. (B2).
Argynnis adippe – 36 ex.: 11.06.2014 – 1 ex. (A), 1 ex. (C); 12.07.2014 – 1 ex. (A), 1 ex. (B2); 04.08.2014 – 3 ex. (A), 1 ex. (B1), 1 ex. (C); 04.09.2014 – 2 ex. (B2), 3 ex. (C); 02.07.2015 – 1 ex. (B1), 6 ex. (B2), 3 ex. (C); 07.08.2015 – 4 ex. (A), 1 ex. (B2), 6 ex. (C); 01.09.2015 – 1 ex. (B1).
Argynnis niobe – 1 ex.: 07.08.2015 – 1 ex. (C).
Issoria lathonia – 3 ex.: 04.09.2014 – 1 ex. (C); 07.08.2015 – 1 ex. (B2), 1 ex. (C).
Brenthis ino – 11 ex.: 11.06.2014 – 6 ex. (B1); 02.07.2015 – 4 ex. (B1), 1 ex. (C).
Brenthis daphne – 4 ex.: 11.06.2014 – 1 ex. (B1), 1 ex. (C); 02.07.2015 – 2 ex. (C).
Boloria euphrosyne – 18 ex.: 04.08.2014 – 14 ex. (A), 3 ex. (B1); 19.05.2015. – 1 ex. (C).
Boloria selene – 94 ex.: 02.05.2014 – 1 ex. (C); 11.06.2014 – 10 ex. (A), 6 ex. (B1), 1 ex. (B2), 7 ex. (C); 04.08.2014 – 1 ex. (B1), 2 ex. (B2); 04.09.2014 – 1 ex. (B2); 06.06.2015 – 2 ex. (B1), 8 ex. (B2), 11 ex. (C); 02.07.2015 – 2 ex. (B1), 2 ex. (C); 07.08.2015 – 14 ex. (A), 10 ex. (B1), 10 ex. (B2), 11 ex. (C); 01.09.2015 – 2 ex. (B1), 2 ex. (B2), 1 ex. (C).
Boloria dia – 15 ex.: 02.05.2014 – 2 ex. (A); 04.08.2014 – 1 ex. (A), 1 ex. (B2); 04.09.2014 – 1 ex. (A), 1 ex. (C); 19.05.205 – 3 ex. (A), 2 ex. (C); 01.09.2015 – 2 ex. (B2), 2 ex. (C).
Vanessa atalanta – 5 ex.: 04.08.2014 – 1 ex. (B2), 1 ex. (C); 04.09.2014 – 2 ex. (C); 07.08.2015 – 1 ex. (C).
Vanessa cardui – 4 ex.: 04.08.2014 – 1 ex. (B1), 1 ex. (B2); 19.05.2015 – 1 ex. (A), 07.08.2015 – 1 ex. (C).
Araschnia levana – 19 ex.: 02.05.2014 – 2 ex. (A); 04.08.2014 – 1 ex. (A), 1 ex. (B1); 04.09.2014 – 2 ex. (B1); 19.05.2015 – 4 ex. (A), 1 ex. (C); 02.07.2015 – 1 ex. (A), 07.08.2015 – 1 ex. (A), 1 ex. (B1), 3 ex. (B2), 1 ex. (C); 01.09.2015 – 1 ex. (A).
Aglais io – 10 ex.: 02.05.2014 – 1 ex. (A); 04.08.2014 – 2 ex. (B1), 1 ex. (B2); 19.05.2015 – 1 ex. (A); 07.08.2015 – 1 ex. (B2); 01.09.2015 – 2 ex. (B1), 1 ex. (C); 07.10.2015 – 1 ex. (A).
Aglais urticae – 13 ex.: 02.05.2014 – 1 ex. (B2); 11.06.2014 – 1 ex. (B1), 1 ex. (C); 04.09.2014 – 1 ex. (C); 06.06.2015 – 2 ex. (B2); 02.07.2015 – 3 ex. (A), 1 ex. (B2); 07.08.2015 – 1 ex. (A), 1 ex. (B1), 1 ex. (C).
Nymphalis antiopa – 2 ex.: 02.05.2014 – 1 ex. (A); 16.04.2015 – 1 ex. (C).
Nymphalis c-album – 11 ex.: 02.05.2014 – 1 ex. (A); 04.08.2014 – 1 ex. (C), 04.09.2014 – 1 ex. (A), 1 ex. (C); 02.07.2015 – 2 ex. (A); 07.08.2015 – 2 ex. (A), 1 ex. (B1); 01.09.2015 – 1 ex. (A); 07.10.2015 – 1 ex. (A).
Apatura ilia – 1 ex.: 02.07.2015 – 1 ex. (C).
Apatura iris – 3 ex.: 12.07.2014 – 1 ex. (B1); 02.07.2015 – 1 ex. (C); 01.09.2015 – 1 ex. (C).
Melitaea phoebe – 1 ex.: 11.06.2014 – 1 ex. (B1).
Melitaea diamina – 12 ex.: 11.06.2014 – 1 ex. (B1), 1 ex. (B2); 06.06.2015 – 1 ex. (B1); 02.07.2015 – 7 ex. (B1), 2 ex. (C).
Melitaea britomartis – 8 ex.: 11.06.2014 – 1 ex. (B1); 02.07.2015 – 1 ex. (B1), 2 ex. (B2), 4 ex. (C).
Melitaea athalia – 74 ex.: 11.06.2014 – 10 ex. (A), 20 ex. (B1), 10 ex. (B2), 8 ex (C); 04.08.2014 – 1 ex. (A), 1 ex. (B2); 04.09.2014 – 1 ex. (B2); 06.06.2015 – 1 ex. (B1), 3 ex. (B2); 02.07.2015 – 1 ex. (A), 3 ex. (B1), 1 ex. (B2), 6 ex. (C); 07.08.2015 – 3 ex. (A), 1 ex. (B2); 01.09.2015 – 2 ex. (B1), 1 ex. (B2), 1 ex. (C).
Pararge aegeria – 8 ex.: 02.05.2014 – 1 ex. (A); 04.08.2014 – 1 ex. (C); 19.05.2015 – 2 ex. (A), 1 ex. (C); 02.07.2015 – 1 ex. (C); 07.08.2015 – 2 ex. (C).
Coenonympha glycerion – 34 ex.: 11.06.2014 – 3 ex. (B1), 3 ex. (C); 12.07.2014 – 10 ex. (C); 02.07.2015 – 9 ex. (B1), 2 ex. (B2), 7 ex. (C).
Coenonympha pamphilus – 131 ex.: 02.05.2014 – 1 ex. (B1); 11.06.2014 – 9 ex. (B1), 1 ex. (B2), 6 ex. (C); 04.08.2014 – 4 ex. (B1), 6 ex. (B2), 1 ex. (C); 04.09.2014 – 2 ex. (B1), 2 ex. (C); 19.05.2015 – 4 ex. (B2), 9 ex. (C); 06.06.2015 – 11 ex. (B1), 15 ex. (B2), 14 ex. (C); 02.07.2015 – 4 ex. (B1), 1 ex. (B2), 5 ex. (C); 07.08.2015 – 1 ex. (A), 7 ex. (B1), 6 ex. (B2), 3 ex. (C); 01.09.2015 – 7 ex. (B1), 6 ex. (B2), 6 ex. (C).
Aphantopus hyperanthus – 52 ex.: 11.06.2014 – 3 ex. (B1), 10 ex. (B2), 1 ex. (C); 12.07.2014 – 5 ex. (B1), 2 ex. (B2), 3 ex. (C); 02.07.2015 – 20 ex. (B1), 4 ex. (B2), 4 ex. (C);
Maniola jurtina – 288 ex.: 11.06.2014 – 2 ex. (B1), 20 ex. (B2), 1 ex (C); 12.07.2014 – 20 ex. (B1), 20 ex. (B2), 15 ex. (C); 04.08.2014 – 3 ex. (A), 3 ex. (B1), 20 ex. (B2), 20 ex. (C); 04.09.2014 – 5 ex. (B1), 4 ex. (C); 02.07.2015 – 28 ex. (B1), 16 ex. (B2), 20 ex. (C); 07.08.2015 – 7 ex. (A), 20 ex. (B1), 35 ex. (B2), 15 ex. (C); 01.09.2015 – 7 ex. (B1), 3 ex. (B2), 4 ex. (C).
Erebia aethiops – 29 ex.: 04.08.2014 – 7 ex. (A), 15 ex. (C); 07.08.2015 – 7 ex. (C).
Erebia medusa – 30 ex.: 06.06.2015 – 9 ex. (B1), 6 ex. (B2), 15 ex. (C).
Melanargia galathea – 54 ex.: 12.07.2014 – 14 ex. (A), 7 ex. (B1), 3 ex. (B2), 9 ex. (C); 04.08.2014 – 1 ex. (A), 1 ex. (B1), 2 ex. (C); 02.07.2015 – 3 ex. (B1), 2 ex. (B2), 3 ex. (C); 07.08.2015 – 3 ex. (A), 3 ex. (B1), 3 ex. (C).
Minois dryas – 45 ex.: 04.08.2014 – 10 ex. (B1), 10 ex. (C); 07.08.2015 – 1 ex. (A), 15 ex. (B1), 2 ex. (B2), 7 ex. (C).
Čanády A, 2016: Contribution to the knowledge of the butterflies (Lepidoptera: Papilionoidea) distribution in surrounding of villages from eastern Slovakia, part 4 – Miroľa (the Laborecká vrchovina Mts). Folia faunistica Slovaca, 21 (1): 73–83.
[in Slovak, with English abstract]
Received 16 December 2015 ~ Accepted 12 May 2016 ~ Published 13 October 2016
© Faunima, Bratislava, 2016

e–ISSN 1336–4529 ISSN 1335–7522


Obrázok 1. Študijné plochy v okolí obce Miroľa.
Čanády A: Butterflies of the Miroľa village (Laborecká vrchovina Mts)
Folia faunistica Slovaca 21 (1) 2016: 73–83
Tabuľka 1. Systematický prehľad odchytených denných motýľov (podľa Pastorálisa et al. 2013) v okolí obce Miroľa na stanovištiach A–Cs.
Biotopová väzba: U: ubikvistický druh; M1: mozofil-1; M2: mezofil-2; M3: mezofil-3; X1: xerotermofil; X2: xerotermofil-2; H: hygrofil; T: tyrfofil motýľov (podľa Beneša et al. 2002). EN – Endangered (ohrozený); VU – Vulnerable (zraniteľný); LC – Least Concern (najmenej ohrozený); NT – Near Threatened (takmer ohrozený); (podľa Kulfana & Kulfana 2001); Mobilita: 1: extrémne sedentárne; 2: veľmi sedentárne; 3: sedentárne 4: skôr sedentárne; 5: menej sedentárne; 6: ochotné rozptýlenia; 7: mobilné; 8: veľmi mobilné; 9: extrémne mobilné (Bartonova et al. 2014).
|
A |
B1 |
B2 |
C |
Σ |
D% |
Biotopová |
Mobilita |
Červený zoznam |
|||||
|
n |
D% |
n |
D% |
n |
D% |
n |
D% |
väzba |
(Slovensko) |
||||
|
Papilionidae |
|||||||||||||
|
Iphiclides podalirius (Linnaeus, 1758) |
3 |
1,3 |
2 |
0,5 |
2 |
0,4 |
7 |
0,4 |
X2 |
4 |
NT |
||
|
Papilio machaon (Linnaeus, 1758) |
2 |
0,9 |
1 |
0,2 |
2 |
0,5 |
5 |
0,3 |
U |
5 |
LC |
||
|
Hesperiidae |
|||||||||||||
|
Erynnis tages (Linnaeus, 1758) |
6 |
1,2 |
3 |
0,7 |
8 |
1,6 |
17 |
1,0 |
X1 |
3 |
LC |
||
|
Pyrgus malvae (Linnaeus, 1758) |
1 |
0,2 |
7 |
1,4 |
8 |
0,5 |
M2 |
3 |
LC |
||||
|
Carterocephalus palaemon (Pallas, 1771) |
5 |
2,2 |
1 |
0,2 |
7 |
1,4 |
13 |
0,8 |
M2, H |
3 |
LC |
||
|
Thymelicus lineola (Ochsenheimer, 1808) |
1 |
0,4 |
16 |
3,2 |
1 |
0,2 |
15 |
3,0 |
33 |
2,0 |
M1 |
4 |
LC |
|
Thymelicus sylvestris (Poda, 1761) |
1 |
0,4 |
30 |
5,9 |
19 |
4,7 |
23 |
4,6 |
73 |
4,4 |
M2 |
3 |
LC |
|
Hesperia comma (Linnaeus, 1758) |
5 |
2,2 |
6 |
1,2 |
3 |
0,7 |
2 |
0,4 |
16 |
1,0 |
X1 |
3 |
LC |
|
Ochlodes sylvanus (Esper, 1777) |
1 |
0,4 |
4 |
0,8 |
3 |
0,7 |
4 |
0,8 |
12 |
0,7 |
U |
4 |
LC |
|
Pieridae |
|||||||||||||
|
Leptidea sinapis (Linnaeus, 1758) |
5 |
2,2 |
12 |
2,4 |
12 |
2,9 |
12 |
2,4 |
41 |
2,5 |
X2, M2 |
6 |
LC |
|
Leptidea juvernica Williams, 1946 |
22 |
4,3 |
19 |
4,7 |
17 |
3,4 |
58 |
3,5 |
H, M2 |
6 |
LC |
||
|
Antocharis cardamines (Linnaeus, 1758) |
2 |
0,9 |
2 |
0,4 |
5 |
1,0 |
9 |
0,5 |
M1 |
4 |
LC |
||
|
Pieris brassicae (Linnaeus, 1758) |
1 |
0,2 |
3 |
0,6 |
4 |
0,2 |
U |
7 |
LC |
||||
|
Pieris rapae (Linnaeus, 1758) |
17 |
7,4 |
15 |
3,0 |
9 |
2,2 |
7 |
1,4 |
48 |
2,9 |
U |
7 |
LC |
|
Pieris napi (Linnaeus, 1758) |
12 |
5,2 |
7 |
1,4 |
5 |
1,2 |
7 |
1,4 |
31 |
1,9 |
U |
7 |
LC |
|
Colias hyale (Linnaeus, 1758) |
2 |
0,9 |
2 |
0,4 |
4 |
1,0 |
1 |
0,2 |
9 |
0,5 |
U |
7 |
LC |
|
Gonepteryx rhamni (Linnaeus, 1758) |
5 |
2,2 |
1 |
0,2 |
5 |
1,2 |
3 |
0,6 |
14 |
0,8 |
M2 |
7 |
LC |
|
Lycaenidae |
|||||||||||||
|
Lycaena phleas (Linnaeus, 1761) |
1 |
0,2 |
1 |
0,2 |
2 |
0,1 |
U |
4 |
LC |
||||
Čanády A: Butterflies of the Miroľa village (Laborecká vrchovina Mts)
Folia faunistica Slovaca 21 (1) 2016: 73–83
Tabuľka 1. Pokračovanie.
|
A |
B1 |
B2 |
C |
Σ |
D% |
Biotopová |
Mobilita |
Červený zoznam |
|||||
|
n |
D% |
n |
D% |
n |
D% |
n |
D% |
väzba |
(Slovensko) |
||||
|
Lycaena dispar (Haworth,1802) |
2 |
0,9 |
7 |
1,4 |
3 |
0,7 |
4 |
0,8 |
16 |
1,0 |
H |
3 |
VU |
|
Lycaena virgaureae (Linnaeus, 1758) |
3 |
1,3 |
7 |
1,4 |
12 |
2,9 |
3 |
0,6 |
25 |
1,5 |
M2 |
4 |
LC |
|
Lycaena tityrus (Poda, 1761) |
3 |
1,3 |
8 |
1,6 |
18 |
4,4 |
6 |
1,2 |
35 |
2,1 |
M1 |
3 |
LC |
|
Lycaena alciphron (Rottemburg, 1775) |
4 |
1,7 |
3 |
0,6 |
9 |
1,8 |
16 |
1,0 |
M1, H |
4 |
VU |
||
|
Lycaena hippothoe (Linnaeus, 1761) |
7 |
3,1 |
30 |
5,9 |
4 |
1,0 |
10 |
2,0 |
51 |
3,1 |
H, M1 |
3 |
LC |
|
Thecla betulae (Linnaeus, 1758) |
1 |
0,2 |
1 |
0,2 |
1 |
0,2 |
3 |
0,2 |
M2, X2 |
2 |
LC |
||
|
Cupido argiades (Pallas, 1771) |
2 |
0,9 |
4 |
0,8 |
16 |
3,9 |
9 |
1,8 |
31 |
1,9 |
X1 |
5 |
LC |
|
Celastrina argiolus (Linnaeus, 1758) |
1 |
0,4 |
1 |
0,1 |
M3 |
5 |
LC |
||||||
|
Phengaris arion (Linnaeus, 1758) |
1 |
0,4 |
5 |
1,0 |
6 |
0,4 |
X1 |
3 |
VU |
||||
|
Plebejus argus (Linnaeus, 1758) |
4 |
1,0 |
10 |
2,0 |
14 |
0,8 |
X1 |
3 |
LC |
||||
|
Polyommatus semiargus (Rottemburg, 1775) |
1 |
0,2 |
1 |
0,2 |
1 |
0,2 |
3 |
0,2 |
M1, H |
4 |
LC |
||
|
Polyommatus icarus (Rottemburg 1775) |
7 |
3,1 |
9 |
1,8 |
5 |
1,2 |
4 |
0,8 |
25 |
1,5 |
U |
3 |
LC |
|
Nymphalidae |
|||||||||||||
|
Argynnis paphia (Linnaeus, 1758) |
1 |
0,4 |
3 |
0,6 |
4 |
0,2 |
M3 |
4 |
LC |
||||
|
Argynnis aglaja (Linnaeus, 1758) |
1 |
0,4 |
1 |
0,2 |
2 |
0,1 |
M2 |
3 |
LC |
||||
|
Argynnis adippe (Denis & Schiffermüller, 1775) |
9 |
3,9 |
3 |
0,6 |
10 |
2,5 |
14 |
2,8 |
36 |
2,2 |
M2 |
4 |
LC |
|
Argynnis niobe (Linnaeus, 1758) |
1 |
0,2 |
1 |
0,1 |
M2 |
3 |
LC |
||||||
|
Issoria lathonia (Linnaeus, 1758) |
1 |
0,2 |
2 |
0,4 |
3 |
0,2 |
U |
7 |
LC |
||||
|
Brenthis ino (Rottemburg, 1775) |
10 |
2,2 |
1 |
0,2 |
11 |
0,7 |
H, M2 |
2 |
VU |
||||
|
Brenthis daphne (Denis & Schiffermüller, 1775) |
1 |
0,2 |
3 |
0,6 |
4 |
0,3 |
X2, M2 |
4 |
LC |
||||
|
Boloria euphrosyne (Linnaeus, 1758) |
14 |
6,1 |
3 |
0,6 |
1 |
0,2 |
18 |
1,1 |
M2 |
4 |
LC |
||
|
Boloria selene (Denis & Schiffermüller, 1775) |
24 |
10,5 |
23 |
4,5 |
24 |
5,6 |
23 |
4,6 |
94 |
5,7 |
M2, T |
3 |
LC |
|
Boloria dia (Linnaeus, 1767) |
7 |
3,1 |
3 |
0,7 |
5 |
1,0 |
15 |
1,0 |
M1, X2 |
5 |
LC |
||
Tabuľka 1. Pokračovanie.
|
A |
B1 |
B2 |
C |
Σ |
D% |
Biotopová |
Mobilita |
Červený zoznam |
|||||
|
n |
D% |
n |
D% |
n |
D% |
n |
D% |
väzba |
(Slovensko) |
||||
|
Vanessa atalanta (Linnaeus, 1758) |
1 |
0,2 |
4 |
0,8 |
5 |
0,3 |
U |
9 |
LC |
||||
|
Vanessa cardui (Linnaeus, 1758) |
1 |
0,4 |
1 |
0,2 |
1 |
0,2 |
1 |
0,2 |
4 |
0,2 |
U |
9 |
LC |
|
Araschnia levana (Linnaeus, 1758) |
10 |
4,4 |
4 |
0,8 |
3 |
0,7 |
2 |
0,4 |
19 |
1,2 |
M2 |
5 |
LC |
|
Aglais io (Linnaeus, 1758) |
3 |
1,3 |
4 |
0,8 |
2 |
0,5 |
1 |
0,2 |
10 |
0,6 |
U |
7 |
LC |
|
Aglais urticae (Linnaeus, 1758) |
4 |
1,7 |
2 |
0,4 |
4 |
1,0 |
3 |
0,6 |
13 |
0,8 |
U |
7 |
LC |
|
Nymphalis antiopa (Linnaeus, 1758) |
1 |
0,4 |
1 |
0,2 |
2 |
0,1 |
M3 |
6 |
LC |
||||
|
Nymphalis c-album (Linnaeus, 1758) |
8 |
3,5 |
1 |
0,2 |
2 |
0,4 |
11 |
0,7 |
M3 |
6 |
LC |
||
|
Apatura ilia (Denis & Schiffermüller, 1775) |
1 |
0,2 |
1 |
0,1 |
M3 |
4 |
LC |
||||||
|
Apatura iris (Linnaeus, 1758) |
1 |
0,2 |
2 |
0,4 |
3 |
0,2 |
M3 |
3 |
LC |
||||
|
Melitaea phoebe (Denis & Schiffermüller, 1775) |
1 |
0,2 |
1 |
0,1 |
X2 |
3 |
VU |
||||||
|
Melitaea diamina (Lang, 1789) |
9 |
1,8 |
1 |
0,2 |
2 |
0,4 |
12 |
0,7 |
T, H |
1 |
VU |
||
|
Melitaea britomartis Assmann, 1847 |
2 |
0,4 |
2 |
0,5 |
4 |
0,8 |
8 |
0,5 |
X2 |
3 |
VU |
||
|
Melitaea athalia (Rottemburg, 1775) |
15 |
6,6 |
26 |
5,1 |
18 |
4,4 |
15 |
3,0 |
74 |
4,5 |
M2 |
3 |
LC |
|
Pararge aegeria (Linnaeus, 1758) |
3 |
1,3 |
5 |
1,0 |
8 |
0,5 |
M3 |
4 |
LC |
||||
|
Coenonympha glycerion (Borkhausen, 1788) |
12 |
0,2 |
2 |
0,5 |
20 |
4,0 |
34 |
2,1 |
X2, H |
2 |
LC |
||
|
Coenonympha pamphilus (Linnaeus, 1758) |
1 |
0,4 |
45 |
8,9 |
39 |
9,6 |
46 |
9,1 |
131 |
7,9 |
U |
3 |
LC |
|
Aphantopus hyperanthus (Linnaeus, 1758) |
28 |
5,5 |
16 |
3,9 |
8 |
1,6 |
52 |
3,2 |
M1 |
3 |
LC |
||
|
Maniola jurtina (Linnaeus, 1758) |
10 |
4,4 |
85 |
16,8 |
114 |
28,0 |
79 |
15,6 |
288 |
17,5 |
U |
4 |
LC |
|
Erebia aethiops (Esper, 1777) |
7 |
3,1 |
22 |
4,4 |
29 |
1,8 |
X2, M2 |
4 |
LC |
||||
|
Erebia medusa (Denis & Schiffermüller, 1775) |
9 |
1,8 |
6 |
1,5 |
15 |
3,0 |
30 |
1,8 |
M2 |
3 |
LC |
||
|
Melanargia galathea (Linnaeus, 1758) |
18 |
7,9 |
14 |
2,8 |
5 |
1,2 |
17 |
3,4 |
54 |
3,3 |
M1 |
5 |
LC |
|
Minois dryas (Scopoli, 1763) |
1 |
0,4 |
25 |
4,9 |
2 |
0,5 |
17 |
3,4 |
45 |
2,7 |
X2 |
3 |
LC |
|
Spolu |
229 |
507 |
407 |
505 |
1 648 |
||||||||
Čanády A: Butterflies of the Miroľa village (Laborecká vrchovina Mts)
Folia faunistica Slovaca 21 (1) 2016: 73–83
Tabuľka 2. Druhová početnosť, početnosť jedincov a hodnoty indexov diverzity a vyrovnanosti spoločenstva motýľov na vybraných študijných plochách (B1, B2 a C) okolia obce Miroľa.
|
Indexy diverzity |
B1 |
B2 |
C |
|
Shannon-Weaverov index diverzity (H) |
3,21 |
2,91 |
3,44 |
|
Shannon-Weaverov index ekvitability (J) |
0,83 |
0,77 |
0,85 |
|
Počet jedincov |
507 |
407 |
505 |
|
Počet druhov |
48 |
43 |
57 |
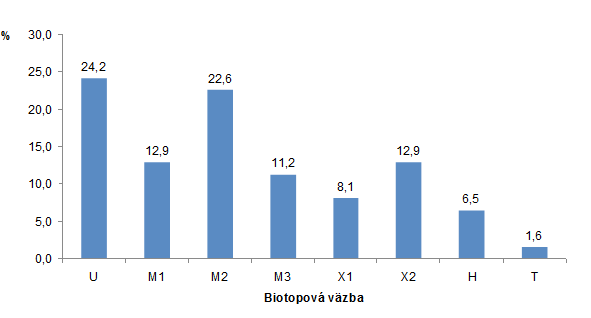
Obrázok 2. Percentuálne zastúpenie denných motýľov okolia obce Miroľa (Laborecká vrchovina) podľa ich biotopovej väzby (BENEŠ et al. 2002): U: ubikvista; M1: mezofil-1; M2: mezofil-2; M3: mezofil-3; X1: xerotermofil-1; X2: xerotermofil-2; H: hygrofil; T: tyrfofil.
Čanády A: Butterflies of the Miroľa village (Laborecká vrchovina Mts)
Folia faunistica Slovaca 21 (1) 2016: 73–83
Čanády A: Butterflies of the Miroľa village (Laborecká vrchovina Mts)
Folia faunistica Slovaca 21 (1) 2016: 73–83